

Year: 2001 Vol. 67 Ed. 5 - (3º)
Artigos Originais
Pages: 618 to 625
Long auditory evoked potential: comparative study between cerebral hemispheres
Author(s):
Ana Claudia F. Frizzo 1,
Renata P. C. Alves 2,
José F. Colafêmina 3
Keywords: long latency auditory evoked potential, event-related evoked potential, cerebral hemispheres
Abstract:
Introduction: Since of the first registration of cerebral electric activities (EEG) in response to the presentation hearing stimulus in human, decade 30's, can be observed the late components or long latency auditory evoked potentials - LLAEP (Mendel, 1989). Ever since, a lot of number of researches have been performed emphasizing the study of the hearing system (the ear and central structures). Considering the functional differences among the cerebral hemispheres, the present study pretend to identify electrophysiologicals evidences to verify differences between hemispheres. Aim: The main objective was to verify the occurrence of possible differences among the captured potentials of the hemispheres right (Cz/A2) and left (Cz/A1) in a group of normal listeners between 8 and 18 years old. Study design: For so much, a comparative analysis of the registrations of LLAEP was performed, as for the latency as the amplitude. Results: Statistical significants differences were not observed among the measures of the derivations Cz/A1 and Cz/A2, except for the component P2 in the masculine population. However, the functional difference between the hemispheres cannot deny, and this should be considered that variable during the performance of the LLAEP's research. Conclusion: Future works will still be necessary, with larger populations or even with different positions of electrodes so that we can verify existence or not of electrophysiologicals evidences that identify those differences, making sure and efficients application of this method.
![]()
INTRODUCTION
Since the 1930's, when the first recordings of cerebral electrical activity (EEG) as a response to auditory stimuli were made in humans, V potentials (vertex), recently known as late potentials or long-latency auditory evoked potentials (LLAEP), could be observed (Mendel, 1989). Since then, many studies have been conducted to emphasize the investigation of the auditory system as a whole (peripheral and central).
However, many advances are still required to make it a very sensitive method for the detection of cerebral abnormalities or defects, since today the cerebral auditory areas that generate late potentials are not clearly defined. Up to the moment, it is known that these potentials are the result of the joint participation of a number of neural structures, that is, multiple generators. Central and parietal regions participate in the generation of responses (Picton & Hillyard, 1974). Lateral neocortex of inferior parietal lobe is responsible for attention and orientation skills of relevant events. There are also simultaneous activities at the hippocampus and frontal lobe. Thus, the inferior parietal lobe interacts with the medial and lateral pre-frontal areas of the attention process and these paralymbic heteromodal areas interact with the hippocampus in the memory processes (Smith et al., 1990).
It is also important to point out that there is wide variability when obtaining and recording the potentials since there is no scientific consensus on the application of the technique, and it is rarely used by audiologists.
It has been known for a long time that there are functional differences between the right and the left hemispheres. After the classical studies by Broca and Wernicke, it was impossible to keep on denying the uneven functional importance of the cerebral hemispheres16. Since then, it is said that right-handed people have speech functions predominantly represented in the left hemisphere. The recognition of verbal auditory stimuli depends on the left hemisphere - and non-verbal auditory stimuli are primarily processed by the right hemisphere (Luria, 1966).
A study comparing stutterers and non-stutterers observed that P300 (a component of LLAEP) of non-stutterers presented statistically significant lower wave amplitudes in the left hemisphere than in the right hemisphere for pure tone stimuli (Kimura, Morgan, Cranford & Burk, 1997). This fact was explained by hemisphere functional asymmetry and higher amplitudes were observed in the right hemisphere, the likely place where non-verbal information (pure tone stimuli) is processed.
It is clear that depending on the stimuli, there is different activation of cerebral auditory areas and we may observe asymmetry in cerebral auditory functioning as a result of the participation of right or left hemispheres.
The research studies on LLAEP have not routinely included the study of wave amplitude because of the frequent oscillation of these values. Nevertheless, this topic has to be considered since by observing it we may functionally compare intra-subject hemispheres (in the same subject), enabling the study of the effects of electrode positioning in the hemispheres and providing better understanding of what is normal and abnormal. The interpretation of amplitude of LLAEP responses will add much to the analysis, leading to further inferences about cortical deficits or lesions.
Taking into consideration the wide application of LLAEP and its inter and intra-subject great variability, we noticed the need to investigate the methodology used in the tests, favoring a safer clinical application of the procedures.
Considering the functional differentiation between the cerebral hemispheres, the present study intended to check the existence of electrophysiological evidence that confirmed inter-hemisphere differentiation. The main objective was to check the presence of differences between potentials recorded for the right hemisphere (Cz/A2) and the left hemisphere (Cz/A1) in a group of normal hearing subjects aged 8 to 18 years. We conducted a comparative analysis of LLAEP recordings considering latency and amplitude, although the current study emphasized amplitude.
Long-latency auditory evoked potentials
LLAEP study has proved to be an effective method to investigate the auditory central nervous system, especially for auditory information processing, since the recording of these potentials reflects the cortical activity involved in brain discrimination, integration and attention skills (Kraus & McGee, 1994).
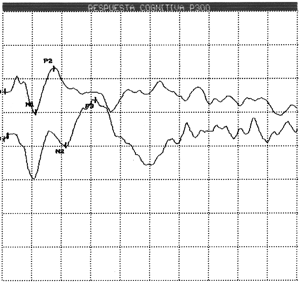
The components of LLAEP are: N1 (P100), P2 (P200), N2 (N200), P3 (P300) (Figure 1).
They are observed 80 to 700ms after the presentation of a given stimulus (McPherson, 1996). They are subdivided into exogenous stimuli (N1, P1, N2), highly influenced by physical characteristics of the stimulus (intensity and frequency, among others) and endogenous potentials (P300), predominantly influenced by internal events related to cognitive skills. P300, cognitive (endogenous) or eventrelated potential, represents the functional use of the stimulus by the subject, with no direct dependence of the physical characteristics of the stimulus.
P300 response is related to the main aspects of mental function: perception and cognition (McPherson, 1996).
Human hearing attention process has been attributed to high central mechanisms (cortical), since attention directed to acoustic stimulus (active attention) seems to cause a significant increase in N1 - P2 components, with no further change to components of short and middle latency potentials (Picton and Hillyard, 1974; Hansen and Hillyard, 1980).
In the process of active attention, response elicitation is a result of the conscious discrimination between two different sound stimuli (tone or voice, one frequent and the other rare/random), that is, there is a response to an unexpected event, named oddball paradigm (Figure 2).
Figure 1. LLAEP tracings of a healthy young adult showing rare and frequent tracings of exogenous (N1, P2, N2) and endogenous (P3). components.
During the test, the attention will be attracted by the rare stimulus, by asking the subject to count it mentally, using his/her fingers or by raising his/her hand. As a result of the attention process, there is a positive wave that takes place about 300ms after the stimulus, with amplitude ranging from 10 to 20 microvolts - v (Sutton, 1965; Polich, 1986).
Rare stimuli may vary in frequency, intensity, clicks in-between speech sounds, bursts in-between clicks and omission of a series of stimuli (Ritter and Vaughan 1969; Ritter et al. 1972; Ford et al. 1973; Picton & Hillyard, 1974).
The intensity of the stimulus used to record LLAEP will also interfere in the collected responses. An increase in intensity of the stimulus will consequently increase amplitude and decrease latency of components N1, P2, N2 and P3 (slightly). It suggests that intensity interferes directly in amplitude and latency of waves (Covington et al., 1996).
It is currently known that a double peak is likely to exist, that is, P300 response would be a combination of two components: P3a and P3 b. According to the studies by Squires et al. (1975), these two components would result from different tasks of active and passive attention that produced different latency and amplitude values. P3a peak would correspond to passive attention processes and P3b would be correlated with active attention processes.
Factors such as gender, time of the day, cognitive skill, personality, type of task, body temperature, season of the year and food intake before the test may contribute to variability of latency and amplitude of P300 (Polich, 1991; Cezar & Munhoz, 1999; Colafemina et al. 2000).
It is also known that eye movements interfere in the recording of cranial electrical activity; however, research studies have shown that asking the subject to refrain from blinking is not a good solution to reduce these movements. It has been shown that such instructions result in amplitude decrease and increase of latency peak at P300 when compared to the same situation without instructions (Ochoa & Polich, 1999).
Variations of P300 values, such as latency increase (Goodin et al., 1978; Polich, 1991) and amplitude decrease (Verleger et al., 1991), as a result of aging have been frequently demonstrated.
Hirayasu et al. (2000), in a study about the effect of gender on P300 changes as a result of aging, the greatest changes were noted in latency of P300 (except amplitude) in male subjects over 30 years of age, compared to same age female matches.
According to the study by Barajas (1990), it has been observed that until the age of 18 years, there is a 19ms/ year decrease of latency as a result of aging, and between 6 and 14 years, the negative correlation is 2.4ms. The decrease of latency resulted from aging is fast and reaches its plateau at 20 years. In the age range from 18 to 78 years, there is increased latency in a positive ratio of 1.25ms/year.
The study of cognitive and neurological disorders of dementia, depression, schizophrenia and Alzheimer's disease has been benefited by the research of P300 (Pfefferbaum et al., 1984; Patterson et al., 1988).
Figure 2. Representation of oddball paradigm.
In general, children as from 5 to 7 years of age present P300 (cognitive potential) with increased latency and decreased amplitude when compared to adults. Wave morphological characteristics and latency and amplitude values reach maturity at adolescence (Musiek, 1989; McPherson, 1996).
Diniz Junior (1996), in a comparative study between children with and without school complaints, showed that long-latency auditory evoked potential presented increased latency values in the groups of children with school problems compared to those without problems, at the same age ranges.
Polich et al. (1990) demonstrated that P300 is influenced by age and cognitive development. The authors also suggested that the measures of long latency potentials in normal children populations may reach an association between age and cognitive maturation.
In children, application of LLAEP encompasses the study of language, learning and perceptual disorders, currently known as central auditory processing disorders.
All these variable should be considered in the investigation and interpretation of potentials in clinical populations in order to avoid mistakes during assessment. Taking into account this concern, we decided to describe the above-mentioned information, despite the fact that it is not directly related to the topic addressed by the present study.
Moreover, a more standardized investigation of LLAEP and the existence of further agreement among researchers will promote the clinical expansion of the method.
Therefore, we believe that the investigation of the occurrence of possible distinctions between recorded evoked potentials of right hemisphere (Cz/A2) and left hemisphere (Cz/A1) may be useful to future analysis of variability during the study of LLAEP.
Cerebral hemispheric functional asymmetry
During centuries, much has been discussed about cerebral morphology and physiology, especially about hemispheric distinction. However, only after the studies by Broca and Wernike was it possible to functionally define the difference between the right and the left hemisphere. According to Luria (1966), after the classical studies by Broca and Wernicke, it was no longer possible to deny the uneven functional importance of the cerebral hemispheres. Since then, it has been inferred that right-handed people have a predominance of speech functions in the left brain.
According to Brandão (1995), the development of spatial orientation and language studied in the central nervous system (CNS) has enabled better understanding of cerebral specialization. The studies confirmed that there are differences between the right and left hemispheres and that it is a exclusively human cerebral functional lateralization. The left hemisphere is essentially specialized in language. The right hemisphere is dominant for spatial visualization and other non-verbal response stimuli.
Kimura (1967) reported that the recognition of verbal auditory stimuli depended on the. left hemisphere and that non-verbal auditory stimuli were processed mainly by the right hemisphere. There is evidence that such differentiation may interfere in the performance of the subject during the conduction of tests that involve auditory perception of different kinds of stimuli. If we take it for granted that stimuli presented to the ear are processed on the opposite ear (contralateral pathway), the verbal stimulation would produce left hemispheric predominance and better right hemispheric perception. The author also stated that there is hemispheric lateralization as time goes by, suggesting that a 4 to 5 year-old child may have speech functions predominantly represented in the left hemisphere. It also showed the possibility of delayed development of hemispheric predominance in male than in female age matches.
MATERIAL AND METHOD
The study counted on the voluntary participation of 34 right-handed subjects - 16 male and 18 female subjects aged 8 to 18 years.
Normal peripheral hearing was a requirement for inclusion in the study in order to prevent interference in potential recordings. We included in the study subjects who had normal hearing confirmed by anamnesis, inspection of external auditory canal and pure tone audiometry with speech discrimination. Subjects' health history was collected and we excluded those who had neurological problems and/ or learning disabilities.
Data collection took place at the Department of Psychobiology, Faculdade de Filosofia Ciências a Letras de Ribeirão Preto - USP. We used secondary data from a research project named Estudo Normativo dos Potenciais Evocados Auditivos de Longa Latência em Crianças de 8 a 17 Anos e Adultos de 45 a 60 anos (Standardized Study on Long Latency Auditory Evoked Potentials in Children aged 8 to 17 years and in Adults aged 45 to 60 years), previously approved by the Ethics Committee on Research, Faculdade de Enfermagem, Ribeirão Preto /SP.
Participants and their legal authorized representatives agreed to take part in the present study by signing an informed Consent Form before data collection.
The study of long-latency evoked potentials (P300) was carried out using the software Ati-Nautilus PE (version 4,19 c - Lermed S.R.L. - Argentina, 1992). Studied parameters were: binaural stimuli (tons burst with 20ms plateau and 5ms rise/fall) in 1,000Hz for the frequent tone and 2,000Hz for the rare tone (probability of 20%); both stimuli at 70dBHL intensity; duration of analysis of 800ms; 0.5 to 30Hz filter; sensitivity of 160 microvolts; alternated polarity.
Derivations Cz/A1 and Cz/A2 were collected from placement of three gold electrodes on the vertex (Cz) and on the right and left earlobes (A2 and A1, respectively), plus grounding electrode on the contralateral ear, following the standard position of the international 10-20 system (jasper, 1958).
Stimuli were transmitted through TDH-39 headsets. We produced 200 stimuli and the software did not record above 160 microvolts ( v).
The test was performed in a semi-dark acoustically and electrically treated room.
For a probability of 20%, it is necessary to present at least 20 rare stimuli to collect stable values of amplitude and latency (Cohen & Polich, 1997). Following such proportion, the number of frequent and rare stimuli presented was enough to prevent wave instability.
Subjects were seated on a slightly declined armchair and were instructed to remain relaxed, but alert to mentally count the rare sounds included in the series of frequent sounds presented.
Skin cleaning was performed with a brush and we used electrolytic paste to optimize electrical conductivity of the electrodes. They were attached with micropore tape. Electrode impedance did not exceed 4khohms (K?).
The test lasted approximately 30 minutes divided into 20 minutes for anamnesis and audiological assessment (criteria for inclusion) and 10 minutes for P300, repeated twice to duplicate responses and assure the reliability of the test.
We studied components N1, P2, N2 and P3. Wave amplitude and latency values were recorded by the computer and printed for further analysis. In order to perform an unbiased analysis, we asked another researcher (an audiologist experienced in electrophysiology) to study the waves.
Amplitude and latency values were recorded by the identification of waves at the maximum amplitude peak, considering the two tracings. For components N1, P2 and N2 we observed both rare and frequent acquisitions and for P300 we considered only the rare tracings.
We performed the statistical analysis One-way Anova (p<0.05*), at the significance level of 5%, to conduct a comparative analysis of latency and amplitude values of derivations Cz/A1 and Cz/A2.
RESULTS AND DISCUSSION
We decided to present at the same time the results and the discussion in order to facilitate understanding of the addressed topics.
Since the study included the analysis of four waves (N1, P1, N2 and P3) and derivations of electrodes Cz/A1 (left hemisphere) and Cz/A2 (right hemisphere), we chose to present the data in arithmetical mean, owing to the fact that a detailed description of all waves and derivations would certainly lead to difficult display of data, impairing understanding.
Data were analyzed comparing the differences between responses from derivations CZ/A1 and Cz/A2 for the whole population of subjects (n=34). Table 1 and Graph 1 show the means found for latency values of components collected from both derivations.
Differences obtained for latency values of derivations Cz/A1 and Cz/A2 for each component were not statistically significant N1 [p=0.534]; P2 [p=0.535]; N2 [p=0.819]; P3 [p=0.479]; similarly, no statistically significant differences were found in other studies relating latency measures of Cz/A1 and Cz/A2 for a group of normal hearing male subjects (Morgan, Cranford & Burk, 1997).
In a study of LLAEP in healthy young adults, they did not observe statistically significant differences between the ears for latency and amplitude values (p>0.05) (Colafemina et al., 2000).
Table 2 and Graph 2 show the means found for amplitude values of the components collected from both derivations.
Differences in amplitude values obtained from derivations CZ/Al and Cz/A2, for each component, were not statistically significant: N1[p=0.411]; P2[p=0.434]; N2[p=0.687]; P3[p=0.701]. Nevertheless, statistically significant differences for amplitude of Cz/A1 and Cz/A2 were found for a group of normal hearing male adults (Morgan, Cranford & Burk, 1997).
The total sample (n=34) was divided and analyzed according to derivation (CZ/A1 and Cz/A2) and gender, since it is currently known that thickness of the scalp and size of head, different in men and women, may interfere in the responses of LLAEP (Polich,1991). Tables 3, 4, 5 and 6 show the mean values of latency and amplitude for the components in both derivations presenting male and female results.
The differences in latency values for derivations Cz/ A1 and Cz/A2 for each component in female subjects were not statistically significant (N1 [p=0.865]; P2 [p=0.544]; N2 [p=0.984]; P3 [p=0.776]). Moreover, differences in latency values for derivations Cz/A1 and Cz/A2 for each component in male subjects were not statistically significant (N1 [p=0.448]; P2 [p=0.845]; N2 [p=0.693]; P3 [p=0,422]).
Differences in amplitude values for derivations Cz/ A1 and Cz/A2 for each component in female subjects were not statistically significant: (N1 [p=0.292]; P2 [p=0.804]; N2 [p=0.354]; P3 [p=0.530]).
Differences in amplitude values for derivations Cz/ A1 and Cz/A2 in male subjects were not statistically significant for components (Nl [p=0.960]; N2 [p=0.664]; P3 [p=0.191]), but they were statistically significant for component P2 [p=0.038*].Table 1. Mean latencies (ms) of LLAEP components according to derivations Cz/A1 and Cz/A2.
Graph 1. Illustration of mean latencies (ms) of components of LLAEP according to derivations Cz/At and Cz/A2.
Table 2. Mean amplitude ( V) of LLAEP components according to derivations Cz/A1 and Cz/A2.
Graph 2. Illustration of mean amplitudes (NV) of LLAEP components according to derivation Cz/A1 and Cz/A2.
Other studies did not find statistically significant differences between the measures of amplitude of derivations Cz/A1 and Cz/A2 for a group of normal hearing male subjects, and the values obtained for derivation A2/Cz were higher than those obtained for A1/Cz (Morgan, Cranford & Burk, 1997).
Conversely, in a study of LLAEP in healthy young subjects there were no statistically significant differences between the ears for latency and amplitude values (p>0.05) (Colafêmina et al., 2000).Table 3. Mean latencies (ms) of LLAEP components according to derivations Cz/A1 and Cz/A2 for female subjects.
Table 4. Mean latency (ms) of LLAEP components according to derivations Cz/A1 and Cz/A2 for male subjects.
Table 5. Mean amplitude ( V) of LLAEP components according to derivations Cz/A1 and Cz/A2 for female subjects.
Table 6. Mean amplitude (p V) of LLAEP components according to derivations Cz/A1 and Cz/A2 for male subjects.
Despite the existence of a functional differentiation between the cerebral hemispheres, LLAEP researchers are not especially concerned about focusing the topic addressed by the present study, that is to say, this is not a variable for investigation of potentials.
The literature shows a scarcity of studies emphasizing the value of derivations collected from the left (Al) and right (A2) hemispheres. It is also a fact that amplitude is not a widely debated topic in LLAEP, leaving room for more frequent discussions on latency.
The wide variation of wave amplitude values result in its use only as a supportive parameter, leading to limited clinical applicability (Munhoz et al., 2000).
Owing to this fact, it was impossible to find a larger number of specific studies about the topic to result in a snore detailed and comparative analysis.
It is important to point out that despite the never ending discussions about functional hemispheric differentiation that have been taking place for centuries, it is still not possible to safely determine the participation of each hemisphere in the skill studied by the procedure.
Nevertheless, we noticed that although there were statistically significant differences only for male subjects in component P2, the great number of variables and the small sample size may justify the absence of inter-hemispheric differences. It is also important to suggest that other electrode positioning conditions should be investigated, promoting the flourishing of knowledge in this area and guaranteeing safe clinical application of these potentials.
CONCLUSION
Considering the initial hypotheses and the results obtained in the investigation, we concluded that despite the fact that there were statistically significant differences only for component P2 in male subjects for derivations CzA1/ CzA2, we should not deny the functional hemispheric differentiation, always taking this variable into account when performing the study of LLAEP. Nevertheless, further studies with larger samples and other electrode derivations are still necessary so that we may check whether there is electrophysiological evidence of the differentiation, ensuring safe and effective application of the method.
REFERENCES
1. AQUINO, A.M.C.M.; JUNQUEIRA, C.A.O.; FRIZZO, A.C.F. Processamento Auditivo Central. In: ALMEIDA, C.R.J. Tratado de Otorrinolaringologia Clinica: otologia. No prelo.
2. BARAJAS, J.J. - The effects of age on human P3 latency. Acta Otolaryngol., 476 (suppl.):157-60, 1991.
3. BRANDAO, M.L. - Psicofisiologia. Rio de Janeiro: Atheneu. 1950p, 1995.
4. CÉSAR, H.A.R. & MUNHOZ, M.S.L. - O P300 auditivo em jovens e adultos saudáveis, com uma nova proposta de resposta: levantar a mão. Acta Awho, 18(1):32-7, jan./mar., 1999.
5. COHEN, J.; POLICH, J. - On the numbers of trials needed for P300. Int J Psychophysiol., 25(3):249-255, 1997.
6. COLAFEMINA, J.F.; DE FELIPPE, A.C.N.; JUNQUEIRA, C.A.O.; FRIZZO, A.C.F. - Potenciais evocados auditivos de longa latência (P300) em adultos jovens saudáveis: um estudo normativo. Rev. Bras. de Otorrinolaringol., 66(2):144-8, 2000.
7. COVINGTON, JW.; POLICH, J. - P300, stimulus intensity, and modality. Eletroenceph Clin Neurophysiol., 100(6):579-84, 1996.
8. DINIZ JUNIOR, J. Contribuição ao estudo do potencial evocado auditivo de longa latência em crianças. São Paulo, 1996. (Dissertação de Mestrado - Escola Paulista de Medicina).
9. FORD, J.M.; ROTH, W.T., DIRKS, S J.; KOPELL, B.S. - Evoked Potential correlates of signal recognition between and within modalities. Science, 181:465-6, 1973.
10. FINLEY, W.W.; FAUX, S.F.; HUTCHESON, J.; AMSTUTZ, L. - Long latency event-related potentials in the evaluation of cognitive function in children. Neurology, 35: 323-7, 1985.
11. GOODIN, S.D.; SQUIRES, C.K.; HENDERSON, H.B.; STARR, A. Age-related variations in evoked potentials to auditory stimuli in normal human subjects. Eletroenceph Clin Neurophysiol., 44:447 458, 1978.
12. HANSEN, J.C. & HILLYARD S.A. - Endogenous brain potentials associated with selective auditory attention. Eletroenceph Clin Neurophysiol., 49:277-90, 1980.
13. HIRAYASU, Y.; SAMURA, M.; OHTA, H., OGURA, C. - Sex effects on rate of change of P300 latency with age. Eletroenceph Clin Neurophysiol, section Clin Neurophysiol., 111(2):187-94, 2000.
14. KIMURA, D. - Functional asymmetry of the brain in dichotic listening. Cortex, 3:163-78, 1967.
15. KRAUS, N. & McGEE, T. - Auditory Event-related Potentials. In: KATZ, J. ed. Handbook of Clinical Auditory. 4Th. ed. Baltimore, Williams & Wilkins, 1994. cap. 27, 406-26.
16. LURIA, A.R. 7 Higher cortical functions in man. 2ª ed. New York, Basic Books, 1966.
17. McPHERSON, D.L. - Late potentials of the auditory system. San Diego: Singular Publishing Group, 147, 1996.
18. MENDEL, M.I. - Potenciais auditivos evocados: potenciais médio e tardio. In: KATZ, J., ed. Tratado de audiologia clínica. 31 ed. São Paulo, Manole, 1989. cap 28, 574-91.
19. MORGAN, M.D.; CRANFORD, J.L.; BURK, K. - P300 event-related potentials in stutterers and non-stutterers. J. Speech Hear. Res., 40:1334-40 (December), 1997.
20. MUNHOZ, M.S.L.; CAOVILLA, H.H.; SILVA, M.L.G.; GANAN~A, M.M.; FRAZZA, M.M. - Potênciais evocados auditivos: aspectos históricos a técnicos In: MUNHOZ, M.S.L., CAOVILLA, H.H., SILVA, M.L.G., GANANCA, M.M. São Paulo: Atheneu, Audiologia clínica. cap. 10, 149-72, 2000.
21. MUSIEK, F.E. - Probing brain function with acoustic stimuli. ASHA, 31:100-6, 1989.
22. OCHOA, C.J.; POLICH, J. - P300 and blink instructions. Eletroenceph Clin Neurophysiol, section Clin Neurophysiol., 111(1):93-8, 2000.
23. PATTERSON, J.V.; MICHALEWSKI, H.J.; STARR, A. - Latency variability of the components of auditory event-related potentials to infrequent stimuli is aging, Alzheimer - Fype dementia, and depression. Eletroenceph Clin Neurophysiol., 71:450-60, 1988.
24. PFEFFERBAUM, A.; WENEGRAT, B.G.; FORD, J.M.; ROTH, W.T.; KOPELL, B.S. - Clinical application of the P3 component of event-related potentials. II. Dementia, Depression and Schizophrenia. Eletroenceph Clin Neurophysiol., 59:104-24, 1984.
25. PICTON, T.W. & HILLYARD, S.A. - Human auditory evoked potentials. II: Effects of attention. Eletroenceph Clin Neurophysiol., 36.191-9, 1974.
26. POLICH, J. - Normal variation of P300 from auditory stimuli. Eletroenceph Clin Neurophysiol. 65: 236-240, 1986.
27. POLICH, J.; LADISH, C.; BURNS, B. - Normal variation of P300 in children: Age, memory span and head size. Int. J Psychophysiol., 9:237-48, 1990.
28. POLICH, J. - P300 in clinical applications: meaning, method and measurement. Am. J. EEG Technol., 31:201-31, 1991.
29. RITTER. W., SIMSON, R., VAUGHAN, Jr. L.D. - Association cortex potentials and reaction time in auditory discrimination. Eletroenceph Clin Neuropbysiol., 33:547-555, 1972.
30. RITTER. W., VAUGHAN, Jr. L.D. - Average evoked responses in vigilance and discrimination a reassessment. Science, 164:326 328, 1969.
31. RUTH, R.A. & LAMBERT, P.R. - Auditory evoked potentials. Otolaryngol. Clin., 24(2):349-370, 1991.
32. SMITH, M.E.; HALGREN, E.; SOKOLIK, M.; BAUDEVA, P.; MUSOLINO, A.; LIEGEOIS-CHAUVEL, C.; CHAUVEL, P. - The intracranial topography of the P3 event-related potential elicited during auditory oddball. Eletroenceph Clin Neurophysiol., 76: 235-248, 1990.
33. SQUIRES, N.K.; SQUIRES, K.C.; HILLYARD, S.A. - Two varieties of long-latency positive waves evoked by unpredictable auditory stimuli in man. Eletroenceph Clin Neuropbysiol., 38:387-401, 1975.
34. SUTTON, S.; BRAREN, M.; ZUBIDO, J.; JOHN, E.R. - Evoked potentials correlates of stimulus uncertainty. Science, 150:1187-8, 1965.
35. VERLERGER, R.; NEUKATER, W.; KOMPF, D.; VIEREGGE, P. - On the reasons for the delay of P300 latency in healthy elderly cubiects. Eletroencenh Clin Neurophysiol.. 79: 488-502. 1991.
1 Speech and Hearing Pathologist, specialized in Audiology at Universidade de Franca-SP; special student at the Department of Psychobiology, Faculdade de Filosofia Ciências a Letras, Ribeirão Preto - USP.
2 Speech and Hearing Pathologist, specialized in Audiology; professor at Universidade de Franca /SP.
3 Ph.D., Professor, Faculdade de Medicina de Ribeirão Preto-USP; Department of Psychobiology, Faculdade de Filosofia Ciências a Letras de Ribeirão Preto - USP.
Address correspondence to: Rua Tenente Catão Roxo, 1573 - Monte Alegre - CEP 14051-140
Ribeirão Preto-SP- Tel.: (55 16) 633 7893 - e-mail: anafrizzo@uol.com.br
Article submitted on March 22, 2001. Article accepted on May 14, 2001.





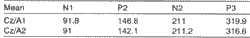

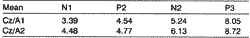

Print: ![]()