A fisiologia do aparelho auditivo é um tema tão amplo que, para uma atualização como a que pretendemos fazer, necessàriamente devemos limitar os assuntos a serem discutidos. Por isso, omitiremos tudo o que se relaciona com a fisiologia do ouvido externo e do médio; para considerarmos sòmente os três pontos seguintes: a) a mecânica da cóclea; b) a eletrofisiologia da cóclea e c) a neurofisiología das vias auditivas.
Mecânica coclear
Para melhor compreender como reage mecânicamente a cóclea quando estimulada por um som, algumas de suas características estruturais devem ser focalizadas. Bàsicamente, a cóclea dos mamíferos é um tubo encaracolado, com duas a quatro voltas, dividido em três partes pelas membranas basilar e de Reissner. Ainda que alguns trabalhos atribuam à curvatura das espiras cocleares um papel importante nas características acústicas do órgão (15) a maioria dos autores considera que o enrolamento assumido é simplesmente uma forma para economizar espaço no osso temporal. Uma outra simplificação que se pode fazer ao analisar a mecânica da cóclea é ignorar a membrana de Reissner, por ser ela acústicamente transparente. Reduz-se, assim, o modêlo coclear a um tubo reto dividido longitudinalmente pela membrana basilar. Ao estudar-se esta membrana, chama a atenção, imediatamente, o fato de sua largura aumentar progressivamente do estribo ao ápice (Fig. 1); além de que, outras características físicas como massa e complacência, também aumentarem continuamente. Estas particularidades determinam o padrão vibratório particular que a cóclea assume ao ser estimulada acusticamente.
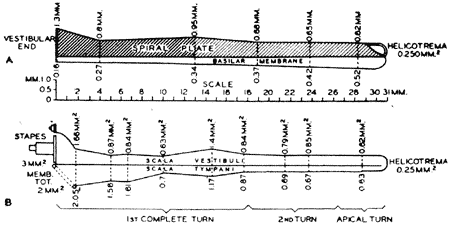
FIG. 1 - A. Forma e dimensão da membrana basilar e da lâmina espiral óssea no homem. B. Diagrama das áreas de secção das rampas vestibular e timpánica. (Da ref. 11.)
O som que passa pelo ouvido externo é transmitido ao ouvido interno através da cadeia ossicular do ouvido médio. O estribo, vibrando na janela oval, atua como um pistão que produz uma onda de pressão na rampa vestibular. Esta onda de pressão, se propaga com a velocidade do som na água (1500 m/s), em vista do que se pode considerar como, pràticamente, a onda atuando símultâneamente sôbre tôda a cóclea, e isso devido às reduzidas dimensões do órgão. De maior complexidade são os efeitos desta onda de pressão sôbre a membrana basilar, em virtude de sua estrutura não ser uniforme. Na espira basal, a membrana basilar é mais leve e rígida, e por isso acompanha fielmente as variações de pressão na rampa vestibular, deformando-se síncrônicamente com as vibrações do estribo. Nos segmentos seguintes da cóclea, onde a membrana possui massa e complacência maiores, as deformações causadas pela onda de pressão e pelas vibrações das porções basais, aparecem com um certo retardo em relação aos movimentos do estribo. Comprovou-se que o retardo aumenta progressivamente com a distância entre os pontos da membrana basílar considerados e o estriba.
O resultado dêsse processo, que pode ser estudado em modelos; e que foi observado diretamente e fotografado em cócleas de cadáveres por von Béhésy (1), é dar origem a uma onda progressiva. Esta pode ser considerada como uma vibração da membrana basilar nascida ao nível do estribo e que se propaga em direção ao helicotrema com uma velocidade comparativamente reduzida e que diminui continuadamente (de 100 a 2 m/s). A amplitude da onda progressiva, aumenta até um determinado ponto e diminui, depois, abruptamente. Dependendo da freqüência do som que estimula, o ponto de máxima amplitude da onda progressiva varia: para freqüências altas se situa na espira basal e para baixas freqüências no ápice. Quando a freqüência é muito baixa (menos de 100 Hz) o comprimento da onda progressiva é grande em relação às dimensões da cóclea, por isso tôda a membrana vibra, pràticamente, em fase.
Resumindo; a) a área da membrana basílar situada próxima ao estribo vibra em resposta aos sons de tôdas as freqüências; b) quanto mais baixa fôr a freqüência tanto maior a extensão da membrana estimulada; c) para freqüências muito baixas tôda a membrana se deforma igualmente.
Deve ressaltar-se que o sentido de propagação da onda progressiva da base ao helicotrema depende das características físicas da membrana e não da posição do estribo. Assim, estimulando-se a cóclea a partir do ápice, ou por vibração de tôda caixa óssea como sucede com os sons que se transmitem por condução óssea, se pode comprovar que o padrão das ondas progressivas não se modifica.
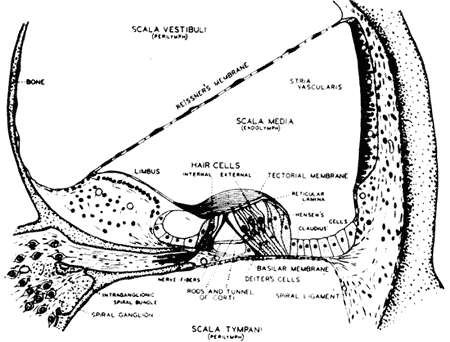
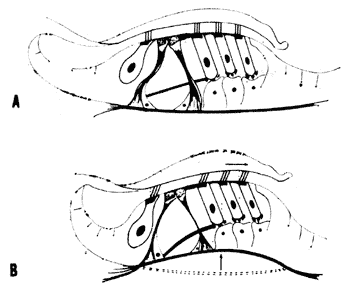
A anatomia da cóclea mostra que o órgão de Corti está firmemente fixado à membrana basilar (Fig. 2) vibrando com ela. As extremidades dos cílios das células receptoras estão presas à membrana tectoria. Como são diferentes os pontos de fixação da membrana basilar e da membrana tectoria, as vibrações do conjunto produzem um deslizamento relativo entre a membrana tectoria e a lâmina reticular, deformando-se, assim, os cílios (Fig. 3). É esta, por conseguinte a forma pela qual as células ciliadas são estimuladas.
FIG. 2 - Diagrama do órgão de Corti e das outras estruturas da rampa média da cóclea da cobaia. (Da ref. 5.)
FIG. 3 - Esquema dos movimento dos cílios causados pela deformação da membrana basilar. (Da ref. 31.)
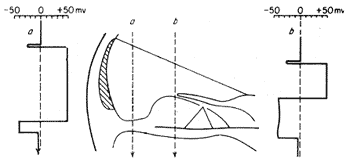
As estruturas que compõem o labirinto apresentam diversos fenômenos elétricos, tanto em repouso como durante a estimulação acústica. Atravessando-se a rampa média da cóclea com um microeletródio, como na experiência de von Békésy ilustrada na figura 4, se constata uma diferença de potencial em relação a um eletródio indiferente, que é negativa, e de cêrca de 60 mV, quando a ponta do eletródio penetra no órgão de Corti; e é positiva, de aproximadamente 80 mV, ao passar para a endolinfa coclear. O potencial negativo mencionado corresponde ao potencial intracelular observado em tôdas as células vivas, e é devido às trocas de ions amplamente estudadas nas fibras nervosas e musculares.
FIG. 4 - Valôres do potencial em diferentes alturas, de corrente contínua medidos por um microeletródio ao atravessar a rampa média. (Da ref. 2.)
Certamente curiosa é a polarização positiva do espaço endolinfático. Foi comprovado que não depende do órgão de Corti, pois que ainda se mantém em animais com degeneração hereditária dêsse órgão. Por outro lado, se modificam profundamente quando o metabolismo da estria vascular é alterado pela administração de tóxicas. O fato de a endolinfa das estruturas vestibulares, com uma composição similar, não apresentar uma polarização apreciável, descarta o papel das altas concentrações do potássio na gênese da polarização positiva. Por isso, êste potencial conhecido oríginàriamente como potencial endolinfático é mais corretamente chamado agora de potencial endococlear. Não existe nada certo quanto à sua função, foi aventada a hipótese que contribuiria para aumentar a sensibilidade das células ciliadas ao incrementar o valor efetivo do seu potencial de membrana. Por outro lado, a comprovação que o valor do potencial endococlear se modifica por deformações da membrana basilar poderia significar que êle intervém no mecanismo gerador do potencial de somação, como veremos mais adíante.
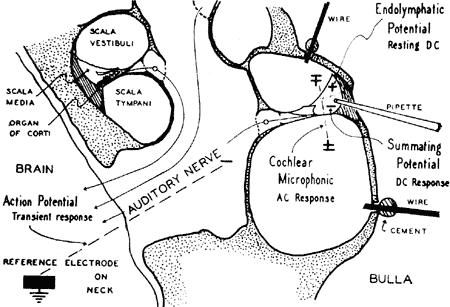
A estimulação sonora da cóclea se acompanha de fenômenos elétricos que podem ser registrados através de eletródios colocados nas proximidades do órgão de Corti. Com esta finalidade, uma disposição muito empregada na cobaia (Fig. 5) consiste em se colocar um par de eletródios, chamados "diferenciais", respectivamente, nas rampas vestibular e timpânica (30) com a que se mede a atividade elétrica do segmento do órgão de Cortí localizado entre êles. Outra técnica, mais simples, emprega um eletródio em contato com a membrana da janela redonda e um eletródío indiferente. Êste segundo método, possibilita registrar, unicamente, a atividade elétrica da espia basal (27); tem a vantagem, no entanto, de permitir o registro dos potenciais cocleares por períodos prolongados e em animais não anestesiados (13, 17).
FIG. 5 - Posição dos eletródios para registro elétrico diferencial na espira basal da cóclea da cobaia. (Da ref. 6.)
Com qualquer tipo de eletródio empregado é possível observar-se um potencial que reproduz fielmente a forma de onda do som usado para estimular o ouvido interno. Por estas características o potencial foi denominado de potencial microfônico coclear. Foi repetidamente demonstrado que êste fenômeno elétrico origina-se ao nível do órgão de Corti, mais especificamente nas células ciliadas. Assim, ao se destruirem estas células, seja mecânicamente ou pela ação de drogas seletivamente tóxicas, como certos antibióticos, ocorre queda no potencial microfônico coclear (Fig, 6), Êsse potencial não tem latência nem limiar apreciável e sua amplitude aumenta linearmente com a intensidade do estímulo até um máximo, decrescendo depois. Tais características são comuns a outros potenciais gradativos dos receptores sensitivos, os potenciais geradores, cuja função é despolarizar as terminações nervosas da via aferente correspondente, iniciando a atividade regenerativa própria das fibras nervosas, e que responde a lei do tudo ou nada.
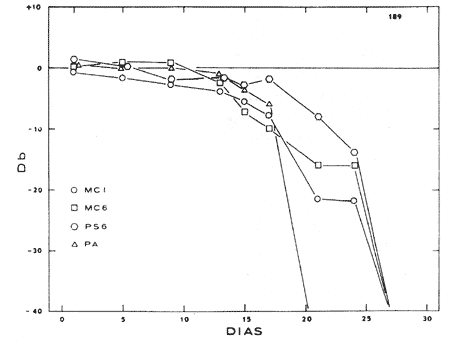
FIG. 6 - Diminuição da atividade elétrica coclear produzida pela administração do antibiótico paromomicina com atividade ototóxica nas doses de 200 mg/kg/dia MC1, MC6: potenciais microfónicos cocleares a 1 e 6 KHz; PS6: potencial de somação positiva a 6 KHz; PA: onda N1 do potencial de ação do VIIIº par. (Da ref. 25.)
Com a técnica dos eletródios diferenciais, foram registrados na cobaia os potenciais microfônicos cocleares nas diferentes espiras cocleares determinados por sons de freqüências variadas. Comprovou-se (Fig. 7), assim, que os sons de elevada freqüência induziam atividade elétrica sòmente na primeira espira, os de freqüência menor na primeira e na segunda e, ùnicamente, os sons de freqüência mais baixa ativavam tôdas as espiras cocleares. Além disso, os registros nas espiras superiores apresentavam um retardo de fase, de caráter progressivo, quando comparados com os potenciais observados na espira basal. Êsses resultados confirmam o padrão de vibração da membrana basilar já descríto, isto é, em forma de onda progressiva.
FIG. 7 - Oscilograma dos potenciais microfônicos cocleares em diferentes freqüências tomadas nas espiras: basal, 2.ª e 4.ª, respectivamente, da cóclea da cobaia. (Da ref. 30.)
Nos registros da atividade elétrica coclear obtidos com amplificadores de faixa ampla, como o da figura 8a, se pode evidenciar um deslocamento da linha de base do potencial microfônico coclear. Fsse deslocamento, que representa a superposição de um outro potencial de oscilação lenta ao microfônico, pode ser individualizado com o auxílio de um filtro passa baixos (Fig. 8b). Éste fenômeno, que Davis (7) designou potencial de somação, varia de polaridade e forma de acôrdo com a freqüência e intensidade do som estimulador; e depende, ainda do lugar onde se efetua o registro, e das condições experientais do animal. Entretanto, a sua origem e o papel que desempenha no processo auditivo são, ainda, desconhecidos. Postulou-se que uma deformação persistente da membrana basilar resultante de vibrações não lineares poderia explicar, em parte a sua gênese (34). Por outro lado existem indícios que mais de um gerador intervém em sua formação, o que explicaria a variabilidade que lhe é característica.
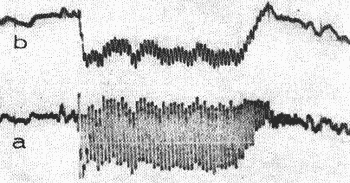
FIG. 8 - A: potencial microfônico coclear induzido por um tom de 700 Hz, 100 db SPL e registrado na janela redonda. b: potencial de somação negativo obtido por filtragem do potencial microfônico a. (Da ref. 18.)
O fato do potencial de somação, aparecer, geralmente com os estímulos que tem intensidade superior àquela necessária para induzir o potencial microfônico coclear, faz pensar que êle interfere na "transdução" auditiva ampliando a faixa dinâmica.
O oitavo par craniano que constituo o primeiro neurônio da via auditiva, tem uma configuração bipolar, cem os corpos celulares no modíolo da cóclea, no gânglio espiral. As suas terminações centrais estão em sinapse com o núcleo coclear e suas ramificações periféricas se distribuem, de forma bastante complexa, sôbre o órgão de Corti (28). Antes de atingir esse órgão, as terminações nervosas atravessam a "habenula perforata" onde perdem a bainha de mielina, dirigindo-se, após, às células ciliadas externas e internas sob a forma de fibras radiais e fibras espiraladas. As células cífiadas externas são inervadas por ambos os tipos de fibras com a particularidade que cada fibra faz sinapse com um número considerável de células (Fig. 9). Nas células ciliadas internas, no entanto, a inervação é preponderantemente feita por fibras radiais que fazem sínapse com algumas poucas células receptoras. Diferentemente das células ciliadas externas as internas recebem fibras de múltiplos neurônios do nervo acústico.
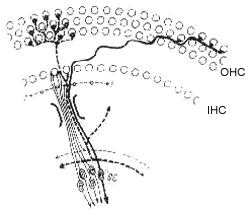
FIG. 9 - Esquema da inervação do órgão de Corti. As fibras aferentes estão representadas em linhas contínuas e as eferentes em pontilhado. As fibras que se dirigem às células ciliadas externas estão em traços grossos e que vão para as ciliadas internas em traços finos. (Da ref. 25.)
Para explicar as particularidades de inervação da cóclea diferentes hipóteses foram aventadas. O fato de múltiplas células cífiadas externas conectarem-se com a mesma fibra nervosa serviria para aumentar a sensibilidade do sistema através do mecanismo de soma espacial, tal como ocorre nos bastonetes retianos. Por outro lado a inervação múltipla das células ciliadas internas concorreria para ampliar a gama dinâmica do receptor, isso se considerarmos que as várias fibras aferentes apresentam limiares distintos. Tanto as células ciliadas externas como as internas possuem uma rica inervação eferente cuja função será discutida mais adiante. As fibras eferentes tem as mesmas particularidades de distribuição que as aferentes. Deve ser destacado que essas fibras fazem sinapse com os corpos das células ciliadas externas, enquanto que nas cífiadas internas a sinapse é feita, aparentemente, só com as fibras aferentes.
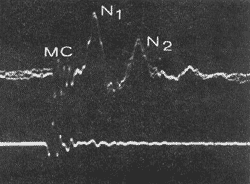
A atividade elétrica de, VIIIº par pode ser registrada através de diferentes métodos. Um dêles, por exemplo, o do eletródio na janela redonda, permite registrar o potencial de ação composto e originado em resposta a estímulo de curta duração ("clic"). Emprega-se êsse tipc de estímulo a fim de que haja sincronização das descargas unitárias de um grande número de fibras do nervo com o que o potencial atinge suficiente amplitude para ser registrado na janela redonda, aparecendo como uma seqüência de duas ou três ondas negativas de amplitude decrescente, com uma latêncía de 0,5 a 1 ms (Fig. 10) a contar do potencial rnícrofónico coclear.
FIG. 10 - Oscilogramas do potencial de ação do VIIIº par (feixe superior), registrado com um eletródio em janela redonda, e do "clic" estimulador (feixe inferior) recolhido com um microfone. MC: potencial microfônico coclear; N1 N2 : ondas do potencial de ação.
Sòmente se observa a amplitude máxima do potencial de ação do nervo em resposta a determinado estímulo na ausência de qualquer outro som. A simultaneidade de outros estímulos, especialmente ruídos com amplo espectro de freqüência (ruído branco), produz uma acentuada queda em tôdas as ondas de potencial de ação, sem modificar, no entanto, o potencial mícrofônico. Êste fenômeno reflete o estado transitório de inexcitabilidade (período refratário) de algumas fibras, determinado pelos potenciais gerados pelos sons, e constitue a base fisiológica para explicar o mascaramento auditivo.
A inscrição de microeletródios no oitavo par permite o registro de fibras isoladas. Isto já foi feito na cobaia, no gato, no macaco e em outros animais (ver para referência (16, 24, 29) observando-se, assim, que há, para cada fibra uma freqüência de estímulo sonoro denominada característica, que induz potenciais nessa fibra com intensidade menor que a necessária para outras freqüências. A medida que se afasta da freqüência característica maior é a intensidade requerida, o que levado em gráfico, onde os valôres de freqüência e intensidade são representados, dá uma curva em forma de V assimétrica. Esta "sintonia" das unidades nervosas do oitavo par decorre da estimulação seletiva de diferentes grupos de células ciliadas em resposta a freqüências distintas, tudo na dependência da característica formação das ondas progressivas na cóclea.
A estimulação auditiva com sons de baixa freqüência (menores que 1 kHz) permite comprovar que algumas unidades do nervo acústico são ativadas ao mesmo tempo que ocorrem as variações de pressão da onda sonora, aparecendo uma espícula em cada ciclo (Fig. 11 ). Êste tipo de atividade nervosa, apesar da limitação imposta pela periodicidade de recuperação das fibras nervosas, é preservada nos relés superiores da via auditiva e permite a análise de freqüência dos tons baixos, em resposta aos quaís todo o órgão de Corti é estimulado e em conseqüência a sintonização das fibras do VIIIº par é pobremente definida. Teríamos, assim, pelas modernas teorias da audição, que a percepção da altura de um som seria feita pelo sistema nervoso central através de um mecanismo duplo: 1.º) a distinção dos sons de alta freqüência dependeria do fato que cada uma delas estimularia grupos de fibras diferentes e isso porque o órgão de Corti seria excitado em uma área relativamente restrita correspondente aos pontos de maior amplitude da onda progressiva. 2.º) para os sons de freqüência baixa, que põem em vibração amplas extensões da membrana basilar, e conseqüentemente ativam um grande número de fibras, a informação quanto à altura seria transmitida pela sincronização das descargas com o som estimulador. A atual teoria dupla combina os dois principais artigos de ressonância localizada de Von Helmholtz e o da teoria telefônica de Rutherford.
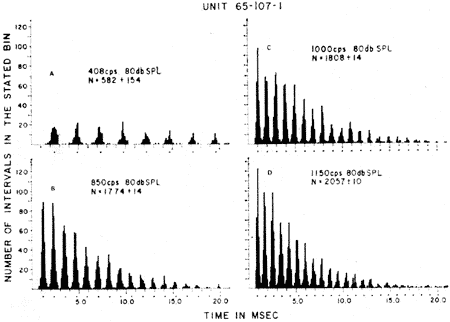
FIG. 11 - Histograma da densidade das descargas unitárias, obtido com um computador digital, mostrando as respostas sincronizadas do nervo auditivo ao estímulo em 4 freqüências diferentes. Duração do estímulo: 1 segundo; número de estímulos computados: 10. (Da ref. 26.)
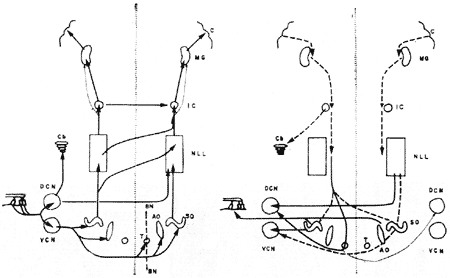
Ao considerarmos as conexões que faz o nervo acústico (Fig. 12), desde logo ressalta o grande número de relés sináptícos, a multiplicidade de vias cruzadas e a extensa rêde de conexões eferentes e isso em todos os nívéis dê sua via.
FIG. 12 - Vias auditivas aferentes (esquerda) e eferentes (direita). DCN: núcleo coclear dorsal; VCN: núcleo coclear ventral; Cb: cerebelo; AO: núcleo olivar acessório; SO: núcleo olivar lateral; IC: colículo inferior; MC: corpo geniculado medial; C: cortex cerebral. (Da ref. 32.)
Tôdas as fibras do nervo acústico fazem sinapse no núcleo coclear, cuja atividade elétrica já foi extensamente investigada. Como o observado no VIIIº par, também neste núcleo cada neurônio apresenta uma freqüência característica nas experiências em que as respostas unitárias são analisadas com microeletródios. O agrupamento de unidades de freqüência próxima em áreas circunscritas é que permite a chamada localização tonotópica. A sintonização das unidades é aí mais acentuada que no nervo; também se observam respostas sincronizadas, porém, com o limite superior de freqüência mais baixo devido às sinapses que foram acrescentadas. Além disso, se constata no núcleo coclear uma atividade de repouso mais intensa; essa atividade, que se recolhe na ausência de estimulação sonora, pode ser inibida em algumas unidades ao se fazer atuar um som.
Ao nível do complexo olivar superior, constituído pelo agrupamento de cinco núcleos, não foi comprovada localização tonotópica. A sintonia ali apresenta características mais agudas e as respostas mais complexas que nos níveis inferiores. Isto, porque além da multiplicidade de sinapses excitatórias e inibitórias que chegam a êsses núcleos também para aí convergem as informações provindas do ouvido contralateral através do corpo trapezóide. Êstes últimos impulsos tanto podem aumentar como diminuir a atividade das células dos núcleos olivares. É a esta altura da via auditiva que se inicia o processo de comparação das informações provenientes de ambos os ouvidos e que finalmente permitirá a localização auditiva. Em gatos a secção do corpo trapezóide determina uma nítida diminuição da capacidade de localizar a origem dos sons (14).
A maioria das fibras originárias do complexo olivar termina no colículo inferior. Neste núcleo as respostas unitárias, induzidas por tons puros, alcançam o máximo de complexidade e diversidade de todo o sistema. As múltiplas interrelações facilítadoras e inibidoras, tanto ípsí como contralaterais, que ocorrem neste nível determinam padrões de atividade elétrica muito variados, taís como: descargas excitadoras puras que se mantém durante todo o estímulo; respostas inibidoras puras que suprimem a atividade expontânea durante todo o estímulo; dois ou três potenciais coincidentes com o início da estimulação seguidos de um curto período de inibição ("período silencioso") que se contínua com uma descarga continuada; atividade inicial de curta duração seguida de descarga persistente; inibição seguida de excitação ao ínterromper-se a estímulação (resposta "off") (15, 16). Diferentemente do complexo olivar o colículo inferior apresenta uma nítida localização tonotópica, como também respostas sincronizadas até 400-500 Hz (Fig. 13).
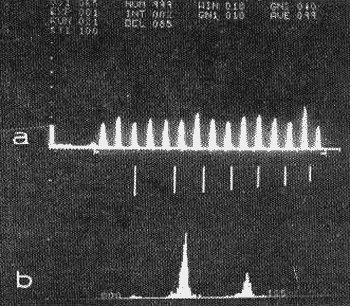
FIG. 13 - Respostas unitárias sincronizadas com o estímulo, no colículo inferior da cobaia. Análise de 100 respostas obtidas com um computador digital. A: histograma de densidade de descargas; B: histograma de distribuição de intervalos entre descargas: Freqüência: 120 Hz; duração: 100 ms. (Da ref. 20.)
Do que acima foi exposto se pode deduzir que na cadeia neuronal da via auditiva, da cóclea ao colículo inferior, a informação sensocial sofre um processamento crescente, representado pela sintonia mais definida das unidades nervosas, pela maior complexidade das respostas, e pela maior participação das informações provenientes do ouvido contralateral. O estudo das estações subseqüentes da via auditiva - corpo geniculado e córtex cerebral - mostra, surpreendentemente, uma inversão dêste processo. Assim, as respostas obtidas no corpo geniculado medial por estímulação auditiva com tons puros são formadas, ùnicamente, por uns poucos potenciais durante os primeiros instantes da estímulação.
A situação é semelhante no córtex cerebral auditivo, onde os neurônios apresentam pouco definidas as freqüências características e escassas respostas à estimulação provocada pelos tons puros. Por outro lado ao se encontrarem numerosas unidades que sòmente respondem a estímulos muito complexos, corno ruídos diversos (18) ou tons modulados em freqüênciar (19).
Foram encontradas unidades que sómente respondem a estímulos crescentes, por exemplo, não respondendo quando a freqüência é decrescente ou é mantida constante. Neste particular o córtex cerebral auditivo se assemelha ao visual, onde Hubel e Wíese1(20) descreveram unidades que unicamente são ativadas quando o movimento das ímagens na retina se faz em determinado sentido.
A participação do córtex cerebral na audição foi, igualmente, investigada através do método de extirpação seletiva de áreas em animais condicionados a discriminar estímulos auditivos, especialmente pelo grupo de Neff (21). Êstes investigadores comprovaram que, no gato, a extirpação bilateral das áreas auditivas corticais não alterava significativamente a capacidade do animal discriminar a altura e a densidade dos tons puros. Entretanto, também notaram que, contràriamente dos animais normais, que ràpidamente aprendem a diferenciar uma seqüência de tons por outra em que a seqüência é diversa (1 kHz - 2 kHz - 1 kHz por outra 2 kHz - 1 kHz - 2 kHz) aqueles desprovidos do córtex auditivo haviam perdido essa capacidade. Outra função auditiva grandemente prejudicada pelas lesões corticais é a da localização auditiva (22).
Para interpretar todos os fatos até aqui mencionados é mister postular a existência de diversos níveis de análise e integração da informação na via auditiva. Ao nível mesencefálico, provàvelmente, se completam os mecanismos que interferem na discriminação tonal simples. As estruturas mais superiores especialmente o córtex cerebral, constituiriam os sistemas de decisão e de integração dos estímulos complexos, da audição direcional e do reconhecimento da linguagem. No entanto, é fundamental ser ressaltado a esta altura que se nem a maioria dos experimentos de eletrofisiología é realizada com tons puros, esta é uma situação artificial, porque na realidade os sons provenientes da natureza são todos acústicamente muito mais complexos. Por conseguinte, é indispensável uma certa dose de prudência ao se postularem os conceitos sôbre a fisiologia da audição.
Paralelamente às vias aferentes descritas, existe todo um sistema de conexões centrífugas que carreia informação das estruturas superiores para as inferiores, e em todos os níveis da via auditiva (Fig. 12). Êste sistema eferente, é similar ao encontrado em outras vias sensitivas e é parte importante no circuito de realimentação que permite que as estruturas centrais exerçam sua regulação sôbre os receptores periféricos. Existem numerosas dessas vias, porém o conhecimento de sua fisiologia ainda é escasso. Uma dessas conexões mereceu particular atenção, e foi objeto de numerosos estudos, trata-se do feixe olívo-coclear de Rassmussen, que se origina ao nível do complexo olivar e termina fazendo sinapses com as células chiadas do órgão de Corti, com um contingente de fibras diretas e outro cruzado. Nas experiências de estimulação do feixe olivo-coclear (23, 24) ocorreu uma redução da amplitude do potencial de ação do nervo acústico e um aumento dos potenciais microfônícos cocleares. No gato verífícou-se que a destruição do feixe parece afetar a discriminação de freqüência (25). No entanto, ainda não é possível no presente precisar-se o real funcionamento do feixe olívo-coclear, e das outras vias menos estudadas na audição normal do homem e dos animais.
Bibliografia1. Von Békésy, G. - Direct observation of the vibrations of the coclear partition under a microscope. Acta Oto-Laryng., 1952, 42:197.
2. Von Békésy, G. - D. C. resting potentials inside the cochlear partition. J. Acoust. Soc. Am., 1952, 24:72.
3. Von Békésy, G. - Experimental modela of the cochlea with and without nerve supply. In: Neural mechanisms of the Auditory and Vestibular Systems, Charles Thomas, Spring fields, 7960:8.
4. Oapps, M. J. - Tesis, University of Illinois, 1967.
5. Davis, H. - Acoustic trauma in the guinea pig. J. Acoust. Soe. Am., 1953, 25:1180.
6. Davis, H. - Initiation of nerve impulses in cochlea and other mechanoreceptors. In Physiological Triggers. American Physiological Society, Washington, D. C., 1957:60.
7. Davis, H., Deotherage, B. H., Eldredge, D. H., Smith, C. A. - Summating potentials of the cochlea. Am. J. Physiol., 1958, 195:257.
8. Destnedt, J. E. - Auditory-evoked potentials from cochlea to cortex as influenced by activation of the efferent olivo-cochlear bundle. J. Acoust. Soc. Am., 1962, 34:1478.
9. Evans, E. F., Whitfield, I. O. - Classificatiou of unit responses in the auditory cortex of the unanesthetized and unrestrained cat. J. Physiol., 1964, 171:476.
10. Fex, J. - Auditory activity in centrifugal and centripetal cochlear fibres in cat. Acta Physiol. Scand., 1962, 55, suppl. 189.
11. Fletcher, H. - Speech and Hearing in Communication, Van Nostrand, Princeton, 1953.
12. Galambos, R. - Mieroelectrode studies ou medial geniculate body of cat. 111. Response to pare tones. ,I. Neurophysiol., 1952, 15: 381.
13. Galambos, R., Roppert, A. - Action of the middle ear museles in normal cat. J, Acoust - Soe, Am., 1959, 31:349.
14. Hubel, D. H., Wiesel, T. N. - Receptor fields, binocular interaction and funetional architecture in the cat's visual cortex. J. Physiol., 1962, 160:106.
15. Huxley, A. F. - Is resonance possible in the cochlea after all? Nature, 1969, 221: 935.
16. Riang, N. Y. S. - Dischange Patterns of Single Fibers in the Cat's Auditorv Nerve. M. I. T. Presa, Cambridge, 1965.
17. Marseillan, R. F. - Estudo da Atividade Elétrica Coclear em Cobaias não Anestesiadas. Metodologia e Aplicações. Tesis, Faculdade de Medicina de Ribeirão Preto, U.S.P., Riberão Prêto, 1963.
18. Marseillan, R. F. - Características do potencial de somação coclear na cobaia não anestesiada. Ciência e Cultura, 1968, 20: 368.
19. Marseillan, R. F. - Mundie, J. R. - Single unit activity in the inferior colliculus of the guinea pig. J. Acoust. Cor. Am., 1965, :17: 1206.
20. Marseillan, R. F., Mundie, J. R. - Respuestas unitaria en el tubérculo cuadrigésimo inferior del cobayo. Acta Physiol. Lat. Amer., 1966, 16, supl. 1:79.
21. Masterton, R. B., Jane, J. A., Diamond, I. T. - Role of brainstem auditory structures in sound localization. J. Neurophysiol., 1967, 30:341.
22. Neff, W. D. - Localization and lateralization of sound in space. In: Hearing Mechanisms in Vertebrates, Churchill, London, 1968:207.
23. Neff, W. D. - Role of the auditory cortex in sound discrimination. In: Neural Mechanisms of the Auditory and Vestibular Systerts, Charles Thomas, Springfield, 1960: 211.
24. Nomoto, M., Suga, N., Katstcki, Y. - Discharge pattern and inhibition of primary auditory nerve fibers in the monkey. J. Neurophysiol., 1964, 27:768,
25. Oliveiro, J. A. A., Marscillan, R. F, - Efeitos da neomicina e da parontomicina sôbre o labirinto anterior da cobaia. Anais XVIII Congres. Bras. Otorrinolaringol., Tomo I, Pôrto Alegre, 1969:201.
26. Rose, J. E., Brugge, J. F., Anderson, D. J., Hind, J. F. - Patterns of activity in single auditory nerve of the squirrel monkey. ht: Hearing Mechanistns in Vertebrates. Churchill, 1968:144.
27. Simmons, F. B., Beattg, D. L. - The significante of round window-recorded cochlear potentials in hearing. An autocorrelated study in the cat. - Ann. Otol. (St. Louis), 1962, 71:767.
28. Spoendlin, H. - Ultrastructure anda peripheral innervation pattern of the receptor in relation to the first coding of the acoustic message. In: Hearing Mechanisms in Vertebrates, Churchill, London, 1968:89.
29. Tasaki, I. - Nerve impulses in individual auditory nerve fibers of guinea pig. J. Neurophysiol., 1954, 17:97.
30. Tasaki, I, Davis, H., Legouix, J, - The space-time pattern of the cohclear microphonics (guinea-pig) as recorded by differential electrodes. J. Acoust. Soc. Am., 1952, 24: 502.
31. Wersäll, J., Mock, A. - Morphological aspects of cochlear hair cell physiology. In Sensorineural Hearing Processes and Disorders, Little, Brown, 1967:3.
32. Whitfield, I. C. - The Auditory Pattiwav. EdxNard Arnold, London, 1967,
33. Whitfield, I. C., Evans, E. F. - Responses of auditory cortical neurons to stimuli of changing frequency J. Neurophysiol., 1965, 28: 655.
34. Whitfield, I. C., Ross, H. F. - Cochlear microphonic and summating potentials and the outputs of individual haircell generators. J. Acoust. Soe. Ant., 1965, 38:126.
* Departamento de Fisiologia. (Lab. de Bioacústica). Faculdade de Medicina de Ribeirão Preto - U. S. P. - Ribeirão Preto - S. Paulo.