INTRODUÇÃOHá uma delicada homeostase entre os capilares do ducto coclear, os fluidos do ouvido interno (perilinfa e endolinfa) e as estruturas que compõem a escala média. Tal equilíbrio depende das trocas de nutrientes e metabolitos entre os capilares e os compartimentos líquidos do ouvido interno. Sabemos hoje que a manutenção de uma composição eletrolítica constante e característica na endolinfa e na perilinfa é essencial para o processo de transdução da energia sonora ao nível do receptor coclear. Neste estudo suspensões de microesferas de poliestireno copolímero (Dow Chemical, Indianopolis-Indiana) foram injetadas no ventrículo esquerdo de chinchillas e em seguida estas microesferas foram demonstradas no interior dos capilares do ducto coclear através de preparações de superfície do ligamento espiral, estria vascularis e lâmina espiral coradas com azul da Prússia. As concentrações de sódio e potássio na endolinfa e na perilinfa de animais que receberam injeções de microêmbolos foram estudadas depois de decorridos diferentes intervalos de tempo a contar da injeção e comparadas com as concentrações normais em chinchillas. Os resultados são apresentados e sua correlação com processos mórbidos humanos usualmente atribuídos a fenômenos vasculares é comentada nesta comunicação.
Alford et al(1) injetou suspenções de microesferas estireno divinil-benzeno nas artérias vertebrais de cães normais e demonstrou mudanças nas funções cocleares e vestibulares através de registros eletrofisiológicos. (eletrococleografia e eletronistagmografia). Para localizar os microêmbolos nos capilares da cóclea eles usaram preparações convencionais de osso temporal: decalcificação, coloração com hematoxilina e eosina e cortes seriados com mícrótomo. Com este método histológico eles demonstraram degeneração segmentar do órgão de Corti, estria vascularis e de neurônios no gânglio espiral. Em dois animais havia hidropsia segmentar da escala média. Este último achado aponta para uma circulação radial da endolinfa e demonstra a produção de hidropsia do ducto coclear a partir de uma lesão capilar.
Igarashi et ai (2) em um estudo similar confirmou os achados acima inclusive o aparecimento de hidropsia segmentar. Foi sua impressão que a grande maioria dos microêmbolos estavam localizados na parede lateral do ducto coclear. Não foi possível demonstrar nenhuma microesfera nos capilares da lâmina espiral usando preparações histológicas convencionais.
Suga et ai (3) usou solução de sulfato de bário para produzir embolização dos capilares cocleares em porcos da guiné. Seus achados são semelhantes aos relatados acima.
A preparação histológica usada nos trabalhos revistos acima não pode fornecer uma distribuição precisa dos microêmbolos nas várias áreas capilares da cóclea porque os vasos são vistos cortados transversalmente e os cortes estão a 100 microns de distância mínima. Para que um microêmbolo seja visto é necessário que o plano de corte coincida com sua localização por acaso. Assim sendo muitas partículas não são vistas nos cortes estudados. As técnicas de preparações de superfície usadas neste estudo mostram todas as áreas capilares da escala média longitudinalmente desde sua porção mais basal até o ápice da cóclea, possibilitando um estudo mais preciso da distribuição dos microêmbolos entre as áreas capilares da parede lateral e da lâmina espiral.
A manutenção de uma composição iônica constante e característica nos compartimentos líquidos que rodeiam o órgão de Corti (endolinfa e perilinfa) é essencial para o processo de transdução da energia sonora no receptor coclear (4). O conhecimento dos mecanismos que mantêm aste ambiente iônico constante é portanto essencial para que possamos compreender os mecanismos etiopatogênicos da surdez neuro-sensorial a nível coclear.
Os capilares da escala média são essenciais para a manutenção da homeostese nos compartimentos líquidos do ouvido interno. Em decorrência disto várias formas de surdez neuro-sensorial no homem (presbicusis, surdez súbita idiopática, surdez acompanhando diabetes mellitus e doenças cardio-vasculares) são atribuídas a distúrbios vasculares no ouvido interno. Estudos de alterações bioquímicas nos fluidos do ouvido interno após a produção de uma lesão capilar pura devem fornecer algum subsídio para o esclarecimento destas formas de surdez
neuro-sensorial.
Nesta comunicação nós descrevemos os resultados obtidos com o estudo das áreas capilares da cóclea em preparações de superfície bem como da composição iônica da perilinfa e endolinfa de chinchillas injetadas com suspensões de microêmbolos no ventrículo esquerdo. A distribuição dos microêmbolos nas várias áreas capilares e as mudanças na composição iônica dos fluidos do ouvido interno em função de tempo são descritas.
MATERIAL E MÉTODOS:Uma suspensão a 0,1% de partículas uniformes de estireno divinilbenzeno* em solução fisiológica foi usada para produzir microembolização. O tamanho das partículas foi escolhido de acordo com Axellsson (5). Este autor mediu os diâmetros dos capilares da cóclea em porcos da guiné, um animal experimental comparável com a chinchilla em porte. A suspensão continha partículas com um diâmetro médio de 5,700 microns e um desvio padrão de 1,500 microns (variação de 7,2 a 4,2 microns) e deste modo não embolizariam vasos maiores, limitando a lesão aos capilares.
Chinchilla normais pesando de 350 a 500 gramas foram anestesiadas com Diabutal* intraperitoneal (0,1cc/100 gramas de peso corporal). 0 pelo do lado esquerdo do tórax foi retirado expondo o pré-córdio de modo a poder visualizar o ictus cordis (choque da ponta) e portanto localizar o ventrículo esquerdo. 0,2cc da suspensão de microêmbolos foram injetados no ventrículo esquerdo usando uma seringa de tuberculina e uma agulha tamanho 25. A posição da agulha no ventrículo esquerdo era confirmada pela aspiração de sangue arterial.
Estudo Morfológico - Os ligamentos espirais, estrias vasculares e lâminas espirais de animais tratados como descrito acima foram estudados em preparações de superfície após perfusão de Azul da Prússia através do coração para encher os capilares. Esta técnica foi descrita por Axellsson (6) e nós a descreveremos suscintamente.
Depois da injeção o tórax é aberto através de uma esternotomia ampla, o pericárdio é aberto e o seio venoso também. O animal sangra pelo seio venoso até o coração parar. Após este evento o ventrículo esquerdo é aberto amputando-se a ponta do coração. Um catéter de polietileno é então introduzido na aorta através do ventrículo esquerdo e seguro em posição por uma ligadura de seda N° 1 em torno do coração. Em seguida inicia-se a perfusão de soro fisiológico com uma pressão hidrostática constante de 70cm de água até que a solução saia pelo seio venoso livre de sangue. Uma solução a 2,5% de Azul da Prússia em água é então perfundida com a mesma pressão hidrostática até que a velocidade de perfusão torna-se muito vagarosa. Os ossos temporais são então removidos e colocados em gluteraldeído a 5 % durante uma noite. A cóclea é então dissecada com o auxílio do microscópio cirúrgico, o ligamento espiral é removido com a estria vascularis em toda a extensão da cóclea a lâmina espiral e o modíolo são removidos in totum e colocados em ácido tricloroacético a 5% para decalcificar. O ligamento espiral e estria vascularis são deshidratados em concentrações ascendentes de álcool etílico, lavados em xilol e montados em lâminas para exame microscópico em sua espessura total. Depois de decalcificados a lâmina espiral é separada do modíolo por microdissecção e montada em lâminas para exame microscópico também em sua espessura total.
Estudos Bioquímicos
Endolinfa e perilinfa foram obtidas por um método desenvolvido no laboratório de Bioquímica do Ouvido Interno do Departamento de Otorrinolaringologia da Faculdade Medicina da Universidade de Minnesota.
Após anestesia. como descrito acima os animais eram decapitados rapidamente e os ossos temporais eram removidos rapidamente e mergulhados em nitrogênio líquido (Temperatura de -150 C). Usando uma câmara de congelação retirada de um sistema para obter secções finas de tecidos congelados** que nos permitia trabalhar na microdissecção, das cócleas sem descongelá-las a seguinte dissecção era feita: as bulas eram abertas e o osso que cobre a cóclea era removido com uma broca (Jordan-Day), o ligamento espiral e a estria vascularis eram removidos usando uma agulha fina e deste modo a cóclea era aberta expondo a perilinfa e a endolinfa congeladas separadamente. Os cristais congelados de endolinfa e perilinfa eram retirados com uma agulha fina separadamente e colocados em lâmina de vidro para liquefazer-se a temperatura ambiente.
Análise das Amostras - As amostras de endolinfa obtidas por esta técnica são da ordem de 5 nanolitros. Para analizar estas amostras um "Helium Glow Photometer"' tem que ser usado. As amostras foram preparadas para análise do seguinte modo: primeiro foram colocadas entre lâminas de óleo mineral em uma pipeta capilar, depois aspirando-as com uma pipeta de 10 nanolitros e colocando-as em 200 nanolitros de Cs-N03-NH4 H2 PO4 como diluente entre lâminas de óleo mineral em uma pipeta capilar selada. Esta pipeta era colocada no "Helium Glow Photometer" e as amostras eram analizadas em alíquotas de 5 nanolitros. As concentrações de sódio e potássio na perilinfa e na endolinfa podem ser determinadas deste modo.
Plano Geral da Investigação: Duas chinchillas (4 cócleas) foram estudadas pela técnica descrita sob o título "Estudo Morfológico" após microembolização numa tentativa de estabelecer a eficiência do método de embolização e de
estabelecer padrões de distribuição dos microêmbolos pelas áreas capilares da cóclea.
Onze chinchillas (22 cócleas) foram usadas para estudos bioquímícos na endolinfa e perilinfa depois da microembolização. A seguinte seqüência de tempo após microembolização foi seguida nos estudos bioquímicos:
- 2 horas após microembolização .......................... 4 animais
- 24 horas após microembolização .......................... 4 animais
- 2 meses após microembolização ......................... 3 animais
Seis animais normais (12 cócleas) foram estudados pelo mesmo método bioquímico para estabelecer padrões normais contra os quais pudéssemos comparar os animais tratados com a suspensão de microêmbolos.
RESULTADOS:As figuras de 1 a 5 mostram microêmbolos no interior de capilares pertencentes a várias áreas capilares da cóclea (estria vascularis, eminência espiral, ligamento espiral por trás da estria vasculares e lâmina espiral). Um número maior de microêmbolos foi registrado nos capilares da estria vascularis e do ligamento espiral do que nos capilares da lâmina espiral. Os capilares da estria continham mais microêmbolos que as demais áreas capilares da parede lateral do ducto coclear. Não foram encontrados microêmbolos nos capilares da lâmina espiral em sua porção mais apical em nenhuma das oito cócleas estudadas morfologicamente.
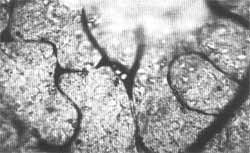
FIG. 1 Estria vascularis de Chinchilla normal após injeção de 0,2 cc de uma suspensão a 0,1 % de partículas uniformes de Estireno Divinilbenzeno através do coração. Técnica de coloração: injeção de azul da Prússia através do coração, 400 X. Seta aponta para partículas dentro de um capilar da estria vascularis.
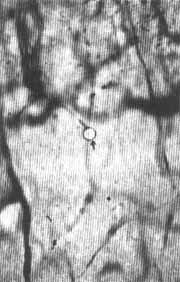
Fig. 2 - Ligamento espiral de Chinchilla normal após injeção de 0,2cc de uma suspensão a 0,1 % de partículas uniformes de Estireno Divinilbenzeno através do coração. Técnica de coloração: injeção de azul da Prússia através do coração, 400X. Seta aponta para partícula dentro de um capilar por trás da estria vascularis.
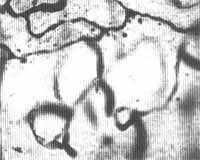
Fig. 3- Ligamento espiral e estria vascularis de Chinchilla normal após injeção de 0,2cc de uma suspensão a 0,1 % de partículas uniformes de Estireno Divinilbenzeno através do coração. Técnica de coloração: injeção de azul da Prússia através do coração, 400X. Seta aponta para partícula dentro de um capilar da Eminência Espiral.
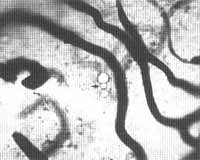
Fig. 4 - Ligamento espiral e estria vascularis do turno apical de Chinchilla normal apos injeção de 0,2cc de uma suspensão a 0,1 % de partículas uniformes de Estireno Divinilbenzeno através do coração. Técnica de coloração: injeção de azul da Prússia através do coração, 400X. Seta aponta para partículas dentro de capilares da estria vascularis. Alguns capilares estão cheios desangue em vez de Azul da Prússia provavelmente devido a obstrução por microêmbolos.

Fig. 5 - Lâmina espiral do turno basal de Chinchilla normal depois de injeção de 0,2cc de uma suspensão a 0,1 % de partículas uniformes de Estireno Divinilbenzeno através do coração. Técnica de coloração: injeção de azul da Prússia através do coração, 400X. Seta aponta para partícula dentro de uma arteriola radial em uma bifurcação.
A Tabela I mostra as concentrações de sódio e potássio na perilinfa e endolinfa depois de vários intervalos de tempo a contar da injeção de microêmbolos através do coração e compara estes valores com os normais. Os resultados do teste estatístico de Student são também indicados e revelam altos níveis de significação estatística para as diferenças encontradas entre animais injetados com micrêmbolos e os normais. (p 0,05) Apenas as concentrações de sódio na endolinfa duas horas após a injeção não foram significativamente diferente das normais. (p 0,2)
TABELA I - Alterações nas Concentrações de Eletrolitos nos Fluidos do Ouvido Interno da Chinchilla em Conseqüência de Microembolizações Capilares.
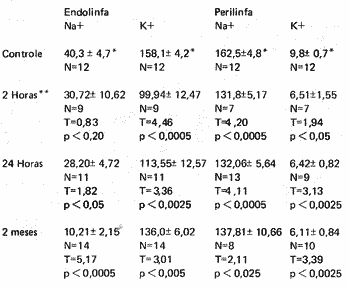
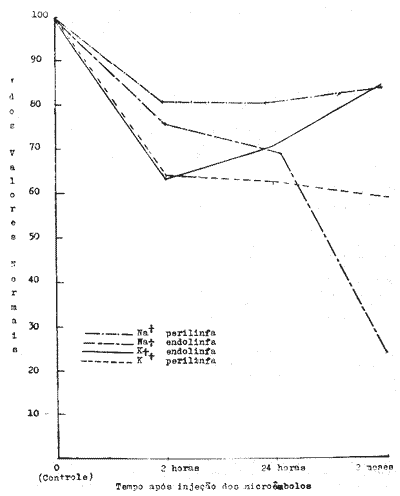
Fig. 6 - Evolução das mudanças nas concentrações eletrolíticas nos fluídos do ouvido interno de Chinchillas em função em função do tempo decorrido desde as injeções de microêmbolos no coração. Concentrações expressas como % dos valores normais (Ver Tabela I).
A figura 6 mostra a evolução das concentrações de sódio e potássio na endolinfa e na perilinfa em função do tempo contado a partir da injeção de microêmbolos. Embora todas as concentrações sejam claramente afetadas, algumas diferenças na evolução das mudanças de concentração entre diferentes eletrólitos no mesmo compartimento líquido e do mesmo eletrólito em diferentes compartimentos líquidos são aparentes. O potássio na endolinfa e o sódio na perilinfa caem rapidamente de concentração e seus valores atingem o nível mínimo duas horas após a injeção de microêmbolos. A partir daí há um retorno lento e incompleto para os valores normais. Estas concentrações estão ainda significativamente abaixo dos valores normais dois meses após a injeção de microêmbolos. As concentrações de sódio na endolinfa e potássio na perilinfa diminuem rapidamente nas primeiras duas horas também mas continuam diminuindo lentamente até os dois meses após a microembolização. O sódio na endolinfa diminue acentuadamente entre 24 horas e dois meses pósmicroembolização atingindo 25% dos valores normais no fim de dois meses. Todas as outras curvas mantêm-se acima de 60% do normal durante todo o período experimental.
Uma lesão aguda ao nível das áreas capilares da cóclea-microembolismo - afeta portanto as concentrações de sódio e potássio na perilinfa e endolinfa e este efeito parece permanente desde que está claramente presente dois meses após a lesão ter sido provocada. Existem diferenças na evolução das mudanças de concentração em função do tempo entre diferentes eletrólitos no mesmo compartimento líquido e entre diferentes compartimentos líquidos em relação ao mesmo eletrólito.
COMENTÁRIOS.:A injeção de microêmbolos na artéria vertebral de animais do porte da Chinchilla é um procedimento difícil. Freqüentemente a dissecção e o manuseio do vaso fazem cessar o fluxo sangüíneo fazendo a injeção de microêmbolos inútil. Além disto o procedimento cirúrgico necessário para expor as artérias em questão é de grande porte e um número significativo de animais não resistem a ele. A técnica descrita de injeção dos microêmbolos através do coração é simples e efetiva em 100% das vezes. Após a recuperação anestésica os animais mostrarão sinais de lesões neurológicas em decorrência de microembolias cerebrais mas eles sobreviverão sem cuidados especiais por longos intervalos de tempo.
A técnica de preparações de superfície usada neste trabalho não foi usada antes em estudos de microembolização.
Ela permitiu uma localização precisa dos microêmbolos nas diversas áreas capilares da cóclea tornando possível o estabelecimento de um padrão de distribuição destes corpos extranhos nos capilares da cóclea. A presença de um número maior de microêmbolos no ligamento espiral e na estria vascularis confirma os achados de lgarashi (2). Especificamente a estria vascularis continha o maior número de microêmbolos no presente estudo.
Os efeitos da microembolização nas concentrações de eletrólitos nos líquidos cocleares não foram descritos na literatura antes. As mudanças acentuadas nas concentrações de eletrólitos na endolinfa e perilinfa descritas acima em conseqüência de lesões nos capilares da cóclea sugerem fortemente que os fluidos cocleares são produzidos ao nível dos capilares da cóclea. A contribuição do líquido céfalo-raquídeo para a formação da perilinfa, se existe, não é importante o suficiente para impedir mudanças permanentes na concentração de eletrólitos neste compartimento líquido após lesões ocorridas nos capilares cocleares.
Embora os microêmbolos tenham sido demonstrados em todas as áreas capilares da cóclea eles estavam presentes em número mais elevado na estria vascularis. Poucos microêmbolos foram encontrados na lâmina espiral dos turnos inferiores da cóclea e nenhum no turno apical. As mudanças em concentrações de eletrólitos podem ser portanto, atribuídas primariamente embora não exclusivamente a lesão de capilares da estria vascularis.
A despeito de não existirem evidências conclusivas a este respeito, acreditase que a função da endolinfa é manter um ambiente iônico apropriado para que ocorram os fenômenos eletroquímicos necessários para a transdução de energia sonora em impulsos nervosos (4). A estria vascularis seria primariamente responsável pela manutenção deste ambiente iônico constante na endolinfa através do transporte ativo de iontes entre a corrente sangüínea e o ducto coclear (7, 8, 9, 10, 11, 12, 13). Além disto Rauch (14, 15) mostrou conclusivamente que há transporte ativo ou pelos menos permeabilidade seletiva para iontes através da membrana de Reissner. Quando ele injetou cloreto de potássio marcado com K42 na escala vestibuli o K42 cruzou rapidamente a membrana de Reissner e foi encontrado na endolinfa mesmo contra o gradiente de concentração. Entretanto quando ele injetou cloreto de sódio marcado com Na24 na escala vestibuli somente traços do isótopo radiativo pode ser encontrado na endolinfa embora o gradiente de concentração fosse favorável à difusão deste ionte. Ele concluiu que potássio pode entrar na endolinfa a partir da perilinfa a uma velocidade 4 a 5 vezes superior a do sódio mesmo contra o gradiente de concentração. Deste modo a membrana de Reissner tem um papel na manutenção do equilíbrio iônico da endolinfa principalmente transferindo seletivamente iontes da perilinfa para a endolinfa.
É dentro desta estrutura de conhecimentos que tentaremos interpretar nossos resultados: logo após o fenômeno embólico o transporte ativo de iontes ao nível dos capilares é afetado, principalmente ao nível da estria vascularis. Isto leva a uma diminuição da concentração de sódio e potássio em ambos os compartimentos mas, como o transporte ativo é necessário principalmente ao nível da endolinfa (a perilinfa tem composição iônica semelhante ao soro), as mudanças mais drásticas ocorrem ao nível da endolinfa. A recuperação do potássio na endolinfa deve ser pelo menos em parte devida a passagem deste ionte seletivamente da perilinfa para a endolinfa através da membrana de Reissner. Isto também explicaria o declínio constante da concentração de potássio na perilinfa. Como o transporte de sódio através da membrana de Reissner é muito mais lento, não é possível manter sua concentração na endolinfa e ela continua a cair. Uma concentração baixa de todos os eletrólitos em ambos os compartimentos reflete uma inabilidade de compensar completamente a lesão vascular. Obviamente as anastomoses capilares extensas ao nível da cóclea devem contribuir para a recuperação. É interessante lembrar que a concentração de potássio na endolinfa é provavelmente responsável pela manutenção do potencial endolinfático de repouso e portanto é vital para o processo de transdução que ocorre no receptor coclear. (4). A concentração de potássio na endolinfa mostra a recuperação mais rápida e mais precoce de todas as concentrações estudadas após uma queda rápida inicial (Figura 6). Mecanismos de homeostasia parecem intervir rapidamente para preservar a função neste caso.
Trabalhos anteriores utilizando microembolização de capilares da cóclea tem consistentemente mostrado a presença de hidropsia endolinfática pelo menos em segmentos do ducto coclear (1, 2, 3). Entretanto o estudo de concentrações eletrolíticas em perilinfa e em endolinfa em casos de doença de Menière têm produzido resultados normais (16, 17). É portanto difícil fazer correlação dos achados neste estudo com hidropsia endolinfática. Estudos de osmolaridade dos fluidos cocleares após microembolização deveriam clarificar este aspecto.
A ocorrência de mudanças pronunciadas na concentração de eletrólitos nos fluidos cocleares em conseqüência de microembolização capilar é uma base bioquímica para a ocorrência de surdez súbita coclear de etiologia vascular. O curso de recuperação descrito acima parece indicar que se a lesão vascular não for extensa o suficiente para causar destruição de estruturas cocleares a audição nestes casos deverá via de regra recuperar-se lentamente.
Finalmente uma palavra de moderação. As Teorias relacionadas com a produção e absorção dos fluidos do ouvido interno são controvertidas e, como foi dito acima, não há evidência conclusiva em favor de nenhuma delas. Nós interpretamos nossos resultados dentro da estrutura de conhecimentos aceitos pela maioria hoje. Interpretações diferentes poderiam ser oferecidas com base em diferentes pontos de vista. O importante é que os dados experimentais aqui apresentados sejam confirmados por outros investigadores. Esta a razão básica para esta comunicação.
SUMMARY:
Suspensions os stirene divinylbenzene homogenous latex particles were injected through the left heart of normal Chinchillas. Following such injections the capillary areas of the Chinchilla cochlea were studied in surface preparations using the Prussian Blue as a staining solution. This way the stirene divinylbenzene beads were demonstrated inside capillaries in ali capillary areas of the cochlea but predominantly in the stria vascularis. In addition the sodium and potassium concentrations were studied in endolymph and perilymph following microembolization and at several time intervals. The results of this experiment are reported and discussed in this paper.
1. Alford, B. R., Shaver, E. F., Rosemberg, J. J., Guilford, F. R.: Physiologic and histopathologic effects of microembolism of the internal auditory artery. Ann Otol Rhinol Laryngol 74:728-748, 1965.
2. Igarashi. M., Alford, B. R.. Konishi, S., Shaver, E. F. GUILFORD; F. R.: Functional and histopathologic correlations after microembolism of -lhe peripheral labyrinthine artery in dogs. Laryngoscope 79:603-623,1969.
3. Suga, F., Preston, J., Snow, J. B.: Experimental microembolization of cochlear vesseis. Arch Otolaryngol 92:213-220, 1970.
4. Lawrence, M-Inner ear Physiology, in Paparella and Shumrick-Otolaryngology, Vol. 1, pages 275298. W. B. Saunders CompanyPhiladelphia, Ist edition, 1973.
5. Axellsson, A.: The vascular anatomy of the cochlea in the guinea pig and in man. Acta Otolaryngologica, Suppl 243, 1968.
6. Axellsson, A., Lipscomb, D.: The vascular pattern of the chinchilla cochlea. Acta Otolaryngologica 79:352-365, 1975.
7. Saxen, A.: Histological studies of endolymph secretion and absortion in the inner ear. Acta Otolaryngologica 40:23-31, 1951.
8. Smith, C. A.: Structure of the stria vascularis and spiral prominence Ann Otol Rhinol and Laryngol 66:521-546, 1957.
9.Engstrom, H. Sjostrand, F. S., Spoendlin, H.: Feinstruktur der stria vascularis beim Meerschweinchen. Parct Otorhin laryngol. 17:69-79, 1955.
10. Kimura, R. S., Schuckneck, H. E.: The ultrastructure of the human stria vascularis. Part I. - Acta Otolaryngologica 69:415-427, 1970.
11. Borghesan, E.: Modality of the cochlear humoral circulation. Lryngoscope 67:1266-1285, 1957.
12. Nomura, Y.: Capillary permeability of the cochlea. Ann Otol Rhinol Laryngol70:81-101, 1961.
13. Ruedi, L.: Some animal experimental findings on the function of the Inner ear. Ann Otol Rhinol Laryngol60:993-1018, 1951.
14. Rauch, S., Kostlin, A.: Biochemische studien horvorgang. Z. Laryng. Rhinol Oto141:56-69, 1962.
15. Rauch, S., Kostlin, A., Schnieder, B., Schnndler, K.: Arguments for the permeability of Reissner membrane. Laryngoscope 73:135-147, 1963.
16. Rauch, S.: Biochemical aspects of the pathogenesis of Menière's disease-Otolaryngologic Clinics of North America October 1968, pages 369-374.
17. Wullstein, H. I., Rauch, S.: Endolymph and perilymph in Menière's disease: a report of findings in tree cases. - Arch Otolaryngol 73:7, 1961.
* Trabalho realizado nos Laboratórios de Bioquímica do Ouvido Interno e de Patologia do osso temporal do Departamento de Otorrinolaringologia da Faculdade de Medicina da Universidade de Minnesota como parte de uma Tese de Doutoramento.
** Professor de Otorrinolaringologia e Coordenador desta Disciplina da Faculdade de Ciências da Saúde da Universidade de Brasília.
* Dow Diagnostics-The Dow Chemical Company-Indianapolis-Indiana
* - Diamond Laboratories Inc. - Des Moines - lowa.
** - Sorvall LTC-2, Newton, Connecticut
* American Instruments Company-Silver Spring, Maryland.
* Média ± desvio padrão
** Tempo decorrido desde a injeção de microembolos no coração N-Número de amostras analizadas T e p são valores do Teste de Student para o nível de significação tica em relação aos valores do controle.
