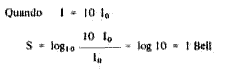Os receptores auditivos estão localizados no sistema auditivo, na sua porção interna chamada cóclea. Na cóclea o órgão de Corti constitui a estrutura receptora. Esta estrutura é mecanoreceptora, respondendo as vibrações sonoras do ar. E um sentido essencial a nossa vida, pois constitui a base da comunicação humana. O ouvido está dividido em três partes: o ouvido externo, o ouvido médio, o ouvido interno. Estudaremos a fisiologia de cada parte separadamente (Fig. 1).
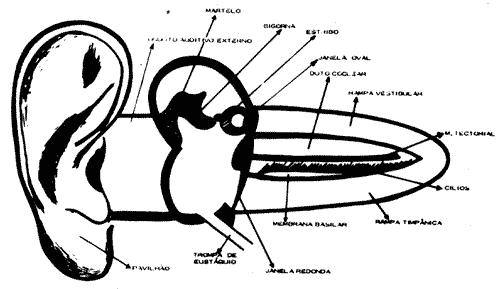
Figura 1 - Estrutura do Sistema Auditivo
OUVIDO EXTERNO
É formado pelo pavilhão e conduto auditivo externo. O pavilhão é formado por cartilagens cobertas por pele, no homem sua importância é discutível, sua finalidade é coletar as ondas sonoras e dirigi-las para o meato auditivo externo. As principais funções deste meato auditivo são: proteção e ressonância sonora. A proteção é exercida pela cera produzida por glândulas ceruminosas na pele do meato. Esta cera forma uma película sobre a pele, que impede a invasão de microrganismos. O meato auditivo externo tem 2,5cm de comprimento e termina na membrana timpânica.
OUVIDO MÉDIO
É a chamada caixa do tímpano, que é limitada externamente pela membrana timpânica e internamente pelo promontório; onde estão as janelas oval e redonda. Superiormente encontramos o ático e antro com células do mastóide e inferiormente a abertura da trompa de Eustáquio. Encontramos no ouvido médio três elementos importantes para a transmissão sonora: a trompa de Eustáquio, os músculos timpânicos e o sistema tímpano-ossicular.
Trompa de Eustáquio - É um canal que comunica a nasofaringe com o ouvido médio, permitindo a entrada de ar nesta cavidade. A ventilação do ouvido médio ocorre durante a deglutição, o espirro, o bocejo, quando a trompa se abre. A trompa permite que as pressões que atuam do lado externo e interno da membrana timpânica sejam igualadas, deixando assim aquela membrana livre para vibrar. Quando a trompa permanece fechada por um resfriado ou variação brusca de altitude, como num avião ou carro subindo ou descendo montanha, sentimos pressão no ouvido, dor e perda de audição e as vezes até ruptura da membrana. Isto ocorre devido a diferença de pressão entre os dois lados da membrana timpânica. Deglutindo com freqüência às vezes conseguimos o desaparecimento dos sintomas. A maioria das infecções do ouvido médio derivam de infecções nasais que se propagaram pela trompa de Eustáquio. Músculos timpánicos - O músculo tensor do tímpano inervado pelo trigêmio (V par craniano) liga-se ao cabo do martelo e quando contrai puxa este ossículo para dentro do ouvido médio.
O músculo do estribo inervado pelo facial (VII par craniano) liga-se à cabeça do estribo e quando se contrai puxa o estribo para dentro do ouvido médio contra a janela oval onde está inserido. Quando o ouvido recebe sons de grande intensidade estes músculos se contraem, aumentando a resistência à transmissão sonora de sons graves Havendo uma queda de aproximadamente 30 dB de intensidade, devido a maior rigidez da cadeia ossicular, protegendo assim o ouvido interno e as células do órgão de Corti contra lesões. A contração reflexa do músculo do estribo pela estimulação sonora com sons intensos é um teste clínico muito usado para a localização de lesões no órgão de Corti através do fenõmeno chamado recrutamento. Além da função protetora que é discutida, este mecanismo mascara sons de banca freqüência em ambientes ruidosos, permitindo a uma pessoa escutar sons de maior freqüência, ou seja, a faixa de freqüência da comunicação falada. Também a contração dos músculos timpãnicos atenua a nossa própria voz quando chega ao ouvido.
Sistema timpano-ossicular - Podemos observar na anatomia comparada do sistema auditivo que todos vertebrados terrestres apresentam ouvido médio e o sistema timpano-ossicular; o que não acontece com os aquáticos, que só apresentam ouvido interno. A presença deste sistema nos animais terrestres foi uma solução evolutiva para adaptação daqueles animais ao meio aéreo. Isto por que o som quando se propaga num meio como o ar, que tem uma impedância própria e passa a se propagar num líquido (ouvido interno) com impedância diferente, perde grande parte da sua energia por reflexão. A membrana timpãnica, martelo e estribo, que formam o sistema timpano-ossicular, são dispostos de tal modo interligados que transmitem as vibrações da membrana timpânica à janela oval e perilinfa da rampa vestibular. É a transmissão aérea. Este sistema timpano-ossicular é um transformador de energia que tem a finalidade de igualar as impedãncias do ouvido médio e interno para uma transmissão sonora efetiva. Este efeito transformador ocorre através de dois mecanismos:
a) Mecanismo hidráulico - A superfície da membrana timpânica tem aproximadamente 55 mm2 e a superfície da janela oval 3,2 mm2. Essa diferença provoca uma multiplicação da pressão sonora, que chega à janela oval, de 17 vezes, o que equivale à razão das superfícies das duas membranas.
b) Mecanismo de alavanca - o martelo e a bigorna atuam conjuntamente constituindo uma alavanca inter-resistente, que tem a potência aplicada no cabo do martelo, sendo a resistência o estribo e o ponto de apoio a articulação da bigorna com o estribo. Este sistema de alavanca, multiplica a pressão sonora, que chega à janela oval aumentada 1,3 vezes e que corresponde a razão entre os comprimentos do martelo e da bigorna.
Os efeitos hidráulicos e de alavanca conjuntamente aumentam a pressão sonora de 22 vezes o que equivale aproximadamente a um aumento de 30 dB, permitindo assim que toda a energia sonora que chegou à membrana timpãnica atinja a base do estribo, perilinfa e órgão de Corti. Uma alteração neste efeito transformador por perfuração da membrana timpãnica ou alteração da cadeia ossicular provocará hipoacusia de transmissão, pois estará alterada a transmissão sonora para o ouvido interno. O sistema tímpanoossicular pode ser reconstituído, quando se
altera na surdez, através de microcirurgia chamada timpanoplastias e pela estapedectomia.
Através de um aparelho, o impedanciõmetro, pode-se medir a impedância do ouvido médio, e pela sua alteração, chegar ao diagnóstico de diferentes distúrbios de transmissão do sistema auditivo. O sistema timpano-ossicular tem
uma freqüência de ressonância de 700 a 1.400 e/s, ou seja, vibra mais facilmente nestas freqüências. O ouvido externo, que apresenta coluna de ar ressonante, tem freqüência ressonante de 3.000 e/s. Daí o nosso ouvido ter uma freqüência ressonante de 600 a 6.000 cps aproximadamente. Além da transmissão chamada aérea, existe no sistema auditivo a condução óssea. Esta ocorre através de vibrações ósseas que atingem diretamente os líquidos do ouvido interno, estimulando os receptores do órgão de Corti. Nossa voz é em parte percebida por nós, por transmissão aérea e em parte por condução óssea. Por isso quando ouvimos nossa voz gravada notamos diferença, pois estamos ouvindo a gravação somente por via aérea. A análise da transmissão aérea de da condução óssea em pessoas suspeitas de surdez constitui a base da audiometria tonal aérea e óssea, que permite o diagnóstico da surdez e o local da alteração. Quando a via aérea está alterada e a óssea normal, temos uma surdez de transmissão e as lesões estarão no ouvido externo ou médio, e se a via óssea estiver alterada haverá hipoacusia neurossensorial, podendo estar afetado o órgão de Corti ou o nervo acústico.
OUVIDO INTERNO - COCLEA
A cóclea ou caracol constitui o labirinto interior que faz parte do ouvido interno ou labirinto. Suas paredes são ósseas, limitando três tubos enrolados em espiral em torno de um osso chamado columela ou modiolo, ao redor do qual dá duas voltas e meia. Para melhor entendermos a anatomia e fisiologia coclear, podemos imaginar a cóclea desenrolada que será vista como na figura 10. Sua base que é mais alargada apresenta as duas janelas, oval e redonda. Os três tubos cocleares têm disposição paralela, e são de cima para baixo: a rampa vestibular que se limita com o ouvido médio pela janela oval; a rampa média ou canal coclear que contém o órgão de Corti; a rampa timpânica que se limita com o ouvido médio pela janela redonda. As duas rampas, vestibular e timpânica, se comunicam pelo helicotrema no ápice da cóclea e contêm perilinfa que apresenta composição química semelhante ao líquido extracelular (maior concentração de íons sódio). O canal coclear apresenta endolinfa com composição semelhante ao líquido intracelular (maior concentração de íons potássio). Como podemos observar na figura 10 a rampa vestibular está limitada do canal coclear pela membrana vestibular de Reissnner e o canal coclear está separado da rampa timpânica pela membrana basilar onde está o órgão de Corti. Esta membrana alarga-se da sua base (0,04mm) até o ápice (0,4mm), aumentando portanto sua massa (Fig. 2).
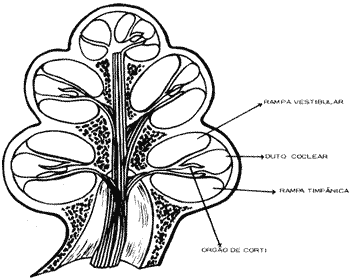
Figura 2 - Estrutura da Cóclea
TRANSMISSÃO DA ONDA SONORA NO OUVIDO INTERNO - ONDAS PROGRESSIVAS (VIAJANTES) - Vom Békesy
O som que é transmitido pelo sistema timpano-ossícular provoca vibrações do estribo que penetram na perilinfa da escala vestibular, ao nível dajanela oval. 0 movimento do estribo para dentro e para fora determina a transmissão da vibração pela perilinfa e esta move-se em direção à rampa timpânica e canal coclear variando a pressão na janela redonda (janela de-descompressão). Durante a transmissão das vibrações pela perilinfa ocorrem deslocamentos simultâneos das membranas do canal coclear, da membrana do Reissnner e da membrana basilar com as vibrações do estribo. Deste modo inicia-se uma onda de oscilação na base da membrana basilar que vai se propagar em direção ao helicotrema até uma certa extensão da mesma, dependendo da freqüência sonora. Estas oscilações da membrana basilar para cima e para baixo têm amplitude em torno de 10-11 mm.
Esta onda propagada "viajante" tem um ponto de deflexão máxima na membrana basilar entre a origem da onda, junto ao estribo, até onde ela termina. Este local de deflexão máxima corresponde à porção da membrana basilar que tem freqüência ressonante natural para a freqüência sonora correspondente. Neste local a membrana vibra com facilidade e a energia da onda dissipa-se. A onda termina neste local. Para cada freqüência sonora, este máximo ocorre em áreas diferentes da membrana basilar. Quanto maior a freqüência, mais próximo do estribo; quanto menor a freqüência, mais próximo do helicotrema. Assim, som agudo provoca uma onda que viaja por uma distância muito pequena na membrana basilar antes de alcançar a sua oscilação máxima e desaparecer. Num som de freqüência média a onda viaja metade da distância antes de desaparecer. Num som grave a onda viaja ao longo de toda a membrana basilar até alcançar a deflexão máxima próxima ao helicotrema. A existência de locais de oscilações máximas localiza cada freqüência sonora num local determinado da membrana basilar.
Neste local as células sensoriais são mais excitadas. Assim cada freqüência provoca excitação máxima em células sensoriais diferentes que informam regiões corticais diferentes podendo ocorrer a discriminação da freqüência sonora (Fig. 3).
DE ALTA FREQUÊNCIA
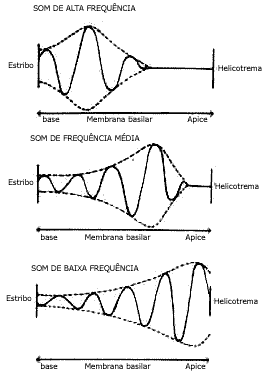
Figura 3 - Propagação da onda vibratória da membrana basilar
ESTÍMULO
Som - é o resultado da transmissão de energia de partículas de ar em vibração, de uma fonte sonora em direção a partes mais distantes.
Freqüência sonora - É expressa em ciclos por segundo ou Hertz. Caracteriza a altura de um som, definindo-o como agudo, médio ou grave, quanto menor a freqüência. A freqüência é inversamente proporcional ao comprimento de onda do som. O chamado tom puro que se usa em audiometria apresenta uma única freqüência. Os sons comuns apresentam várias freqüências.
As freqüências da fala são de 300 c/s a 3.000 c/s.
Intensidade sonora - Corresponde à amplitude das vibrações periódicas das partículas de ar e está associada a pressão e energia sonora (som fraco e som forte). É expressa em dinas por centímetro quadrado ou por decibéis.
Conceito de Bell e decibel - Considerando que o limiar de audição para a freqüência de 1.000 c/s é de 2.10-4 d/cm' no sistema CGS, este é o nível de pressão sonora.
Sendo 1o a energia do som-padrão (limiar) podemos comparar a intensidade de qualquer som com este som-padrão, aplicando a fórmula:
OB = sensação sonora provocada pelo som-padrão, que corresponde ao limiar auditivo na freqüência de 1.000 c/s.
1 Bell é a sensação sonora provocada por um som de energia 10 vezes maior que a do som-padrão. Usa-se o decibel para indicar a sensação de intensidade sonora porque o Bell é uma unidade grande.
Para calcular diretamente o número de decibéis usa-se:
ESTRUTURA DOS RECEPTORES - ÓRGÃO DE CORTI
É a estrutura receptora auditiva, formada basicamente por células de sustentação e células receptaras secundárias ciliadas (estereocílios). Estas células estão situadas sobre a membrana basilar em toda a sua extensão. Temos dois tipos de células ciliadas: uma fileira de células internas (3.500), três fileiras de externas (20.000). Acima do órgão de Corti existe uma membrana gelatinosa chamada tectorial, presa à lâmina espiral óssea do modiolo. Esta membrana cobre os cílios das células ciliadas, entrando em íntimo contato com eles durante as vibrações da membrana basilar. Nos pólos inferiores das células ciliadas encontramos sinapses com neurônios que apresentam seus corpos celulares nos gãnglios espirais de Corti, localizados na cóclea. Os axõnios destes neurônios constituem o nervo coclear. O fornecimento de energia para a cóclea e a manutenção da composição dos líquidos cocleares é realizado pela "stria vasculares", região onde se concentram vasos sangüíneos.
Uma técnica que vem sendo muito utilizada atualmente para estudo da estrutura histológica do órgão de Corti é a técnica de preparações de superfície, na qual o órgão de Corti como um todo é analisado com microscopia de contraste de fases ou microscopia eletrônica de varredura. Deste modo as lesões provocadas em animais como cobaias e coelhos podem ser estudadas, quando estes animais são submetidos a ruídos intensos ou administração de drogas ototóxicas.
MECANISMO DE ESTIMULAÇÃO - TRANSDUÇÃO NO RECEPTOR (Fig. 4)
Como vimos anteriormente, o som determina a oscilação da membrana basilar alternadamente em direção à rampa superior e inferior da cóclea.
A membrana tectorial vai se deslocar com padrão diferente da membrana basilar. Estes deslocamentos da membrana tectorial e basilar, determinarão o aparecimento de forças tangenciais sobre os cílios das células ciliadas, provocando inclinação dos mesmos, pois estes estão em contato com a membrana tectorial.
Podemos registrar com microeletrodos os potenciais existentes na cóclea e chamados potenciais de repouso.
Potencial endococlear é a diferença de potencial existente entre a endolínfa e a perilinfa e de valor de + 80 mV, com positividade na parte interna do canal coclear e negatividade na rampa vestibular. Acredita-se que este potencial seja gerado pela secreção de potássio pela estria vascular no canal coclear.
Papel do potencial endococlear - A endolinfa banha os extremos superiores das células ciliadas e a perilinfa os inferiores. Como as células ciliadas . têm um potencial de membrana de -70 mV em relação à perilinfa, terão um potencial de -150 mV em relação à endolinfa, na superfície superior junto aos cílios. Este elevado potencial elétrico torna a sensibilidade dos receptores muito grande, capaz de despolarizar suas membranas com pequenos movimentos ciliares. Quando a membrana basilar se desloca para cima, os cílios se inclinam e ocorre despolarização dos receptores, isto porque ocorreu alterações de permeabilidade da membrana destes receptores e conseqüente entrada de sódio, despolarização, potencial receptor, neste caso estes potenciais geradores formados determinarão uma freqüência de descarga crescente de impulsos nervosos (potenciais de ação) nas fibras do nervo coclear. Quando a membrana se desloca para baixo, ocorre hiperpolarização e diminuição de freqüência de impulsos nas fibras correspondentes do nervo coclear. Como vimos, a vibração sonora da membrana basilar provoca uma voltagem alternante de mesma freqüência que se superpõe ao potencial de repouso endococlear. Esta voltagem alternante (despolarização, hiperpolarização), constitui os microfônicos cocleares. Estes potenciais se comportam como microfones, reproduzindo as flutuações da pressão sonora. Representaria a soma dos potenciais receptores das células ciliadas excitadas. Suas características são: ausência de período refratário, ausência de latência, ausência de fadiga, limiar não demonstrável.

Figura 4 - Mecanismo de estímulação do órgão de Corti
Para provocar o aparecimento dos potenciais citados e dos potenciais de citados e dos potenciais de ação do nervo acústico, usa-se um estímulo sonoro que é o click Pode-se assim com eletrodos junto à janela redonda registrar estes potenciais no homem e em animais, constituindo as bases neurofisiológicas da eletrococleografta, muito usadas em crianças, para medir os limiares auditivos, orientando a prescrição de prótese e reabilitação da comunicação oral. Em animais, estes registros são usados por exemplo para se estudar as alterações auditivas provocadas pela administração de drogas ototóxicas, principalmente antibióticos aminoglicosídios, como akanamicina, agaramicina, a amicacina, a tobramicina, a estreptomicina, a neomicina e outros.
Limiar auditivo - corresponde a pressão sonora mínima na qual um som numa dada freqüência torna-se audível. Existe um limiar diferente para cada freqüência sonora, sendo os menores limiares encontrados entre 2.800 e 4.000 Hz, ou seja, nestas freqüências a sensibilidade auditiva é máxima. A medida que a intensidade da pressão sonora aumenta, a sensação percebida aumenta, até que o indivíduo sinta dor, alcançando o limiar doloroso, sendo que esta pressão pode danificar o ouvido se a exposição for prolongada.
Faixa de audibilidade - as freqüências audíveis vão de 16 c/s a 20.000 c/s. Acima de 20.000 c/s são os ultra-sons e abaixo de 16 c/s, os infra-sons. A determinação clínica do limiar auditivo é a audiometria, fazendo-se um audiograma com o aparelho chamado audiômetro. Com este exame por via aérea e óssea, chega-se à localização da lesão no sistema auditivo.
Limiar de discriminação de intensidade é a menor diferença perceptível entre dois sons de intensidades diferentes e mesma freqüência. Em baixas intensidades é de 1 dB, podendo ser menor em intensidades maiores.
Limiar de discriminação de freqüência é a menor diferença perceptível entre dois sons de mesma intensidade. Em torno de 1.000 c/s é de 3 Hz. Adaptação auditiva é o aumento do limiar auditivo a um determinado som. Deve-se aos receptores periféricos e neurônios centrais.
VIAS AUDITIVAS (Fig. 5)
As fibras aferentes primárias que vêm do gânglio espiral do órgão de Corti chegam ao núcleococlear ventral e dorsal, fazendo-se sinapse nestes núcleos do bulbo. Do núcleo coclear ventral, fibras nervosas dirigem-se para o complexo olivar dos dois lados. O complexo olivar de cada lado recebe fibras dos dois ouvidos. Neste nível portanto já pode ocorrer a comparação das características sonoras que chegam aos dois ouvidos, para detecção da direção do som. Do núcleo coclear ventral partem fibras que se dirigem para o lado oposto fazendo sinapse no núcleo do leminisco lateral. As fibras que saem do complexo olivar seguem em direção superior, umas do mesmo lado e outras cruzando para o lado oposto, ligando-se aos núcleos do leminisco lateral. Existem fibras que cruzam para o lado oposto neste nível. Em seguida as vias auditivas passam pelo colículo inferior e também cruzam para o lado oposto. Do colículo inferior as fibras passam ao corpo geniculado medial e daí ao córtex auditivo no lobo temporal. Estes tratos são formados por cinco ou seis neurônios. Na cadeia neuronal das vias auditivas da cóclea ao colículo inferior a informação sensorial sofre um processamento crescente, pela sintonia das unidades nervosas, pela maior complexidade das respostas e pela maior participação de informações do ouvido contralateral. No corpo geniculado medial as respostas a estímulos sonoros com tons puros são representadas por poucos potenciais durante os primeiros instantes de estimulação. No córtex aparecem poucas respostas aos tons puros e mais a ruídos complexos. As vias auditivas mantêm conexões com o sistema reticular ativador, cerebelo.
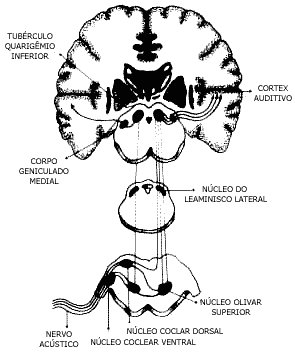
Figura 5 - Vias auditivas
Os potenciais de ação formados nas vias auditivas podem ser registrados, colocando-se um eletrodo ativo no vértex craniano, um eletrodo de referência no meato auditivo externo e um terra na fronte. Deste modo, com uma aparelhagem de estimulação, amplificação, promediação e registro de respostas elétricas, obtém-se sete ondas que corresponderiam às diferentes estações das vias auditivas. Este tipo de registro tem grande aplicação em neurologia e otoneurologia para localização de lesões nas vias auditivas, bem como para medir os limiares audiométricos de crianças por ser um teste objetivo. É a chamada audiometria de respostas elétricas de tronco cerebral.
Vias eferentes - De cada nível do sistema nervoso central, desde o córtex até a cóclea, partem fibras eferentes que chegam ao órgão de Corti. Estes feixes nervosos são inibitórios, reduzindo a sensibilidade dos receptores do órgão de Corti.
Este mecanismo é importante para focalização da atenção em determinados sons, excluindo outros. Mascaramento - Os potenciais de ação do nervo acústico, somente aparecem com amplitude máxima em resposta a um determinado estímulo, na ausência de outro som. A aplicação de ruídos simultâneos, por exemplo o ruído branco, com amplo espectro de freqüência, produz queda acentuada em todas as ondas do potencial de ação, sem modificar o potencial microfônico coclear. Isto se explica pela ocupação de todas as fibras do nervo acústico pelos potenciais provocados pelo ruído branco. O estímulo apresentado certamente ocorrerá nos períodos refratários das fibras não aparecendo potenciais. Isto é a base do mascaramento usado em audiometria clínica.
Localização sonora - Para a localização da direção da fonte sonora é importante a audição com os dois ouvidos. O som alcançará o ouvido mais próximo primeiro e com intensidade maior, isto porque a velocidade do som no ar é constante, e percorrendo espaços diferentes chegará em tempos diferentes nos dois ouvidos. O ouvido humano pode captar diferenças de 3.10-5 segundos. O núcleo acessório da oliva superior é a sede mais importante dos processos neuronais dos impulsos originados do dois ouvidos e que determina a orientação espacial. A análise das diferenças são transmitidas a níveis superiores e ao córtex.
REFERÉNCIAS BIBLIOGRÁFICAS1. BÉKÉSY, G. V. Current status of theories of hearing. Science, 1956,123:779.
2. CASE, J. Sensory Mechanisms. London, Collier-McMillan Limited, 1967.
3. CHERUBINO, M. L'orechio nella senescenza. Giorn. Geront., 1977, 25:88-98.
4. DAVIS, H. Biophysics and physiology of the inner ear. Physiol. Rev., 1957, 37:1.
5. ELDREDGE, D.H. and MILLER, J.D. Physiology of hearing. Ann. Rev. Physiol., 1972, 52:237.
6. GIBSON, W.P.R.; RUBEM, R.J. Essentials of Clinical Electric Response Audiometry.
7. KEIDEL, W.F.; NEFF, W.D. Handbook of sensory physiology. Vol. V-1, V-2, V-3, Berlin, Heidelberg, New York, Springer, 1975.
8. LOWESTEIN, O. The senses. Inglaterra: Chaucer Press Ltda. 1966.
9. LUDEL, J. Introduction to sensory processes. San Francisco: W.H. Freeman and Company, 1978.
10. MARSEILLAN, R.F. Fisiologia coclear. Revista Brasileira de Otorrinolaringologia, 1970, 36-17.
11. MARSEILLAN, R.F.; OLIVEIRA, J.A.A. Critérios de normalidade na análise da resposta elétrica do tronco cerebral humano (BERA) Revista Brasileira de Otorrinolaringologia, 1979, 45:234.
12. MARSEILLAN, R.F.; OLIVEIRA, J.A.A., Del Vechio, F. - Audiometria de respostas elétricas do
tronco cerebral humano. BERA Revista Brasileira de Otorrinolaringologia, 1977, 43:229.
13. OLIVEIRA, J.A.A. Efeitos do salicilato de sódio do labirinto da cobaia. Tese de Doutoramento. Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo, 1971, 95pp.
14. OLIVEIRA, J.A.A. Anatomofisiologia animal comparada. Ribeirão Preto. Editora Universitária, 1971, 408pp.
15. OLIVEIRA, J.A.A. Citoarquitetura coclear. Revista Brasileira de Otorrinolaringologia, 1973,39:12.
16. OLIVEIRA, J.A.A. Sistema coclear. Estimulação das células ciliadas. Revista Brasileira de Otorrinolaringologia, 1974, 40:4.
17. OLIVEIRA, J.A.A., MARSEILLAN, R.F. Estudo das lesões cocleares pela técnica de preparações de superfície na cobaia. Revista Brasileira de Otorrinolaringologia, 1971, 37:1-5.
18. STARR, A. Neurophysiological mecanisms of sound localization. Fed. Proc. 1974,33:1911.
19. STEVENS, S.S., WARSHOFSKY, F. Som e Audição. Rio de Janeiro: Livraria José Olympio Editora, 1968.
20. VAN BERGEIJK, W.A. et al. Waves and the Ear. Garden City, N.Y. Doubledey, 1960.
21. WELLS, C.E.C. Disordes of the special senses in the older patient. Pract, 1975, 215:539-599.
22. WEVER E.G. Electrical potencials of the cochlea. Physiol. Rev. 1966, 46:102.
23. WEVER, E.G. and Lawrence, M. Physiological Acoustics. Princeton: Princeton University Press, 1954.
24 WOLSTENHOLME, G.E.W.; KNIGHT, J. Sensorineural Hearing Loss. London: J. & A. Churvhill, 1970.
* Professor Adjunto Laboratório de Bioacústica Professor Ricardo F. Marseillan - Faculdade de Medicina - Ribeirão Preto - USP. Laboratório de Biocomunicação - Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto - USP. Departamento de Oftalmo-Otorrinolaringologia - Faculdade de Medicina de Ribeirão Preto - USP.