Embriologia da cocleaNo embrião de 18 dias mais ou menos 7 somitos formam um espessamento no ectoderma superficial, bilateralmente ao rombencéfalo (Fig. 1). Estes espessamentos são as placodas auditivas ou óticas. Experimentam a seguir uma rápida invaginação (Fig. 2) formando as vesículas auditivas ou óticas (otocistos) entre 25-30 dias (Fig. 3). Continuando o desenvolvimento, na quinta semana a vesícula auditiva se divide em duas porções, a saber; um componente ventral que originará o século e o duto coclear que inclui o órgão de Corti e um componente dorsal que originará o utrículo, canais semi-circulares e conduto endolinfático (Fig. 4). As estruturas epiteliais formadas desta maneira recebem o nome genérico de labirinto membranoso. Nas etapas iniciais, estas estruturas tubulares estão incluidas no mesênquima. Com o tempo o tecido conjuntivo laxo do mesênquima se converte em um córtex cartilaginoso ou pericondrio que se ossifica e forma o labirinto ósseo. Assim o labirinto membranoso fica incluído por completo no ósseo e ambas as formações estão separadas só por espaços perilinfáticos de pequeno calibre. Durante a formação da vesícula auditiva, separa-se de sua parede um pequeno grupo de células que forma o gânglio estato-acústico. Este gânglio ulteriormente se diferencia em porção coclear e vestibular que guardam relação com as células sensitivas do órgão de Corti e com o sáculo, utrículo e canais semicirculares respectivamente. Os prolongamentos próximos do gânglio coclear e vestibular unem-se para formar o nervo estatoacústico (VIII) que se dirige ao cérebro.
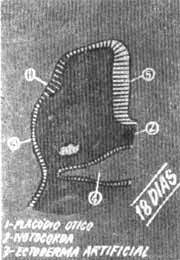
FIG. 1 - Corte transversal esquemático na altura do rombencéfalo num embrião de 18 dias.
4- faringe - 5 - rombencéfalo.
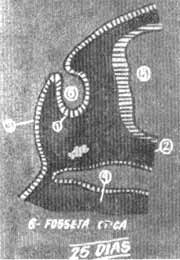
FIG. 2 - Corte transversal esquemático na altura do anterior num embrião de 25 dias.

FIG. 3 - Corte ao mesmo nível que os anteriores no embrião de 5 semanas.
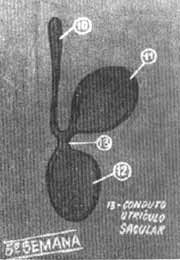
FIG. 4 - Esquema otocisto se dividindo durante a 5.ª semana. 10- conduto endolinfático. 11- utriculo. 12- sáculo.
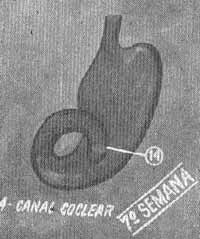
FIG. 5 - Sáculo e canal coclear na 7.ª semana.
Formação da coclea e órgão de Corti
Na sexta semana do desenvolvimento a porção sacular da vesícula auditiva continua sua diferenciação e forma uma evaginação tubular em sua parte inferior (Fig. 5). Este broto, que é o conduto coclear, se introduz no mesênquima circundante formando uma espiral até no final da 8.a semana (Fig. 6). Nestas circunstâncias, sua comunicação com a porção restante do sáculo se resume em um conduto estreito - o canal ou conduto de Hensen.
Pouco depois o mesênquima que rodeia o conduto coclear se transforma por diferenciação em duas porções: uma membrana basal delgada e fibrosa que reveste o exterior do canal e um cortex volumoso de cartilagem. Na décima semana, este cortex cartilaginoso começa a experimentar dissolução (Fig. 7), de modo que se formam dois espaços perilinfáticos, a rampa vestibular e a rampa timpânica (Fig. 8). Nestas circunstâncias o duto coclear fica separado da rampa vestibular pela membrana vestibular ou de Reissner e da rampa timpânica pela membrana basilar. A parede lateral do duto coclear segue unida à cartilagem adjacente em virtude do ligamento espiral; seu ângulo interno está unido e parcialmente sustentado por um prolongamento cartilaginoso longo, que formará futuro eixo do caracol ósseo onde se formará a lâmina espiral óssea. Na etapa inicial, as células epiteliais do conduto coclear são semelhantes. Ao continuar o desenvolvimento, algumas delas se diferenciam e formam duas cristas; a maior, situada perto do centro, se chama crista interna (no adulto a crista sulcada). Esta crista forma a fileira de células ciliadas internas e 3 fileiras de células ciliadas externas ou seja as células sensitivas da audição. Estas células estão cobertas pela membrana tectorial (Fig. 9) que, na etapa inicial, é uma substância gelatinosa e fibrilar que leva consigo a crista sulcada e cuja extremidade se apoia sobre as células ciliadas. O conjunto das células neuroepiteliais e a membrana tectorial que as cobre constituem o órgão de Corti, o órgão verdadeiro da audição. Os impulsos que recebe são transmitidos ao sistema nervoso central pelas fibras auditivas do 8.º par craniano.
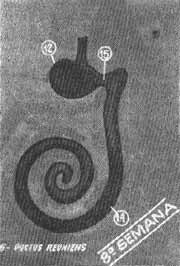
FIG. 6 - Conduto coclear desenvolvido na 8.ª semana.
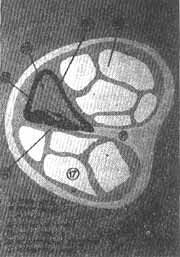
FIG. 7 - Secção esquemática do canal coclear ósseo.
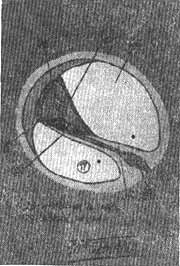
FIG. 8 - Secção semelhante a anterior mostrando as duas rampas vestibular e timpânica do canal coclear ósseo e o canal coclear membranoso.
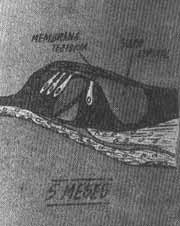
FIG. 9 - Esquema do órgão de Corti e visão transversal.
Estrutura da coclea: (Fig. 10)
A coclea humana e de outros mamíferos vista em secção transversal em um plano que passa pelo eixo do modíolo, aparece como um canal ósseo circular enrolado em um pilar axial (o modíolo). 0 conjunto todo forma aproximadamente um cone, sendo este maior na base e diminuindo rapidamente em direção ao ápice, onde as espiras opostas tornam-se quase contíguas. O próprio pilar (modíolo) tem também aproximadamente a forma cônica. Pela forma do conjunto, conseqüentemente o canal coclear descreve da base para o ápice uma espiral com o raio de curvatura que se torna gradativamente menor. O diâmetro do canal coclear é maior na espira basal. Gradualmente se reduz a 1/4, 1/3, e mesmo 1/2 na espira apical. A espessura da parede óssea que fica entre as espiras também varia, sendo maior entre a espira basal e média enquanto fica reduzida a uma placa fina entre a espira média e apical. O modelo é cheio de cavidades e canais que constituem passagem para os nervos e vasos sanguíneos. A cavidade imediatamente próxima ao limite interno do cana! coclear é o canal do gânglio espiral.
Lâmina espiral óssea, escala vestibular e timpânica do canal coclear
Da parede interna (modiolar) de cada espira do canal coclear nasce a lâmina espiral óssea. Com as partes não ósseas divide o canal em dois compartimentos ou galerias; o superior chamado escala ou rampa vestibular e uma rampa inferior ou timpânica. A largura desta lâmina óssea decresce marcadamente da base ao ápice. Quando vista em secção transversal, a margem livre externa ou lateral da lâmina espiral óssea aparece mais espessa do que sua raiz, ou a extremidade interna, onde é ligada ao modíolo. A margem externa, reforçada e ampliada por tecido conjuntivo forma o limbus que é longitudinalmente percorrido pelo sulco espiral interno. Este corre ao longo de toda a lâmina separando o lábio vestibular superior do inferior ou timpânico. Cada lábio tem suas relações anatômicas peculiares e funções. O lábio vestibular firmemente edificado, também chamado "habenula sulcata" se estende lateralmente em uma estrutura achatada de aparência homogênea; a membrana tectorial de Claudius, também conhecida como membrana de Corti. Em uma secção transversal a membrana tectorial parece uma estreita saliência pendente ao longo do comprimento do duto coclear e com um grande número de fibras delgadas. A parte interna próxima ao modíolo, é ligada ao lábio vestibular da lâmina espiral óssea: a outra margem olhando para a periferia da coclea, fica livre no espaço coclear. Funciona como uma fita elástica capaz de movimentos ondulados ou vibrações em sua extremidade externa.
Lâmina espiral membranosa
O lábio timpânico ou inferior é também chamado "habenula perforata" devido a uma fileira de aberturas pequenas através das quais feixes de fibras nervosas cocleares passam as suas estações terminais. Este lábio aprofunda-se às vezes mais lateralmente no canal coclear do que o lábio vestibular. Em direção a sua extremidade ele torna-se mais fino. Em vez de terminar com uma margem livre, continua como membrana basilar de Claudius, que é lisa. Quando examinada de cima ou de baixo, a membrana basilar parece ser composta de um grande número de fibras lisas e retas. As fibras basilares são dispostas paralelamente uma às outras alterando seu curso suavemente de modo que elas permanecem, em todos segmentos da coclea, radialmente ao eixo modiolar. As extremidades periféricas das cordas basilares, ou da membrana basilar, são fixas à parede externa ou periférica do canal coclear ósseo, embora não diretamente; em vez disto, justamente no ponto de ligação da membrana basilar o plano interno do canal torna-se consideravelmente mais espesso, formando o ligamento espiral de Koelliker. Em secção transversal o ligamento espiral tem a aparência de um crescente. Sua estrutura interna, revela numerosas células e fibras, da natureza do tecido conjuntivo que limita as cavidades ósseas. Cordões fibrosos são encontrados atravessando todas as direções, com a maioria, contudo, convergindo para um ponto - a crista ou margem interna, do ligamento espiral, precisamente onde a membrana basilar é fixada. Neste ponto as fibras juntas do ligamento espiral constituem um tecido compacto que aparece quase homogêneo, dando a impressão de estrutura ligamentosa rígida cujo papel é servir como uma linha de ligação da membrana basilar, aos lábios timpânicos da lâmina espiral óssea. Com a membrana basilar e ligamento espiral fechando o espaço entre a lâmina espiral óssea e a parede externa do canal coclear ósseo a partição do canal em duas escalas é completa. É somente no alto da coclea, onde a partição óssea membranosa não alcança inteiramente o fundo cego do canal, que as duas escalas intercomunicam-se através do helicotrema.
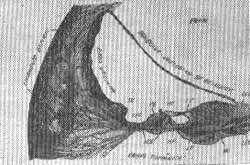
FIG. 10 - Corte esquemático transversal da cóclea.
PE - Proeminência espiral
SEE - Sulco espiral externo
PB - Papila basilar
MB - Membrana basilar
MT - Membrana tectorial
HP - Habenula perforate
LV - Lábio vestibular da Leo
LT - Lábio timpânico da Leo
NC - Nervo coclear
LLE - Limbus da lâmina espiral
LEO - Lâmina espiral óssea
Escala intermediária ou duto coclear
O compartimento superior do canal coclear não ocupa todo o espaço localizado acima da divisão óssea membranosa. Uma membrana muito fina e flexível composta de uma camada de células epiteliais planas e algumas fibras estende-se inclinada de um ponto na superfície vestibular da lâmina espiral óssea, até a parede externa do canal coclear, que é limitado pelo ligamento espiral. Esta membrana, a membrana vestibular de Reissner, com a maior divisão óssea membranosa e com a porção do ligamento espiral, delimita um espaço ou canal que em secção transversal tem a forma de um triângulo estreito: o duto coclear membranoso, ou escala intermediária. É a extensão coclear do labirinto membranoso. Como este duto coclear termina em fundo cego em suas extremidades timpânica e apical a endolinfa que o preenche não tem comunicação com a perilinfa contida nas escalas vestibular e timpânica. Fisicamente, contudo, supõe-se que o duto coclea, estando no mesmo compartimento vestibular, pode atuar como uma parte da escala vestibular. Do sáculo do qual filo e autogeneticamente deriva, o canal coclear torna-se separado no desenvolvimento pela obliteração do ducto de conecção, duto "Reuniens" de Hensen. A escala vestibular e timpânica, ao contrário, permaneceu em comunicação através do aqueduto coclear, com o espaço subaracnoideo do cérebro, com o qual é homólogo. O duto coclear, ou escala intermediária é a parte mais importante da coclea. Contém o analisador psicológico dos sons e também as estruturas por meios das quais o som físico, através de instrumentabilidade do analisador provoca as reações nervosas que são subjetivamente interpretadas no cérebro como sensações auditivas.
Membrana basilar e papila basilar ou órgão de Corti: (Fig. 11)
É a membrana que, devido a sua estrutura e propriedades físicas, é capaz de movimentos oscilatórios. Está estendida entre dois pontos fixos aos quais as fibras da membrana são ligadas; para dentro, o lábio timpânico da lâmina espiral óssea, e para fora a crista interna do ligamento espiral. É constituída de acordo com Retzius, (1884) de aproximadamente 24.000 fibras individuais. As fibras estão dispostas paralelamente em ângulo reto ao comprimento da membrana. Nela são distintas 2 zonas, uma interna ou zona arquada, zona que vai do lábio timpânico à base dos pilares externos e uma zona pectinada entre os pilares externos e o ligamento espiral. No homem todas as fibras da membrana basilar são arranjadas em uma fileira; em alguns mamíferos por exemplo gato ou coelho, há 2 planos de fibras, um acima do outro. O lado inferior da membrana basilar é limitado por um plano irregular de células. Dentro dele, abaixo do túnel interno, há usualmente um vaso sanguíneo, o vaso espiral; ocasionalmente existem vários vasos sanguíneos. Também naquela face estão os feixes nervosos, passando através dos anéis da lâmina espiral óssea para a "habênula perforata". O lado superior ou vestibular da membrana basilar, de face para o duto coclear, apresenta, em contraste, uma superestrutura chamada, o órgão de Corti. Este é o órgão que se supõe específico da audição. O órgão de Corti aparece como uma porção espessada e elaborada do epitélio que limita o lado de dentro do duto coclear.
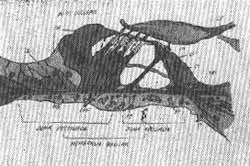
FIG. 11 - órgão de Corti. Vista transversal.
Estrutura suporte do órgão de Corti ou papila basilar
A estrutura do órgão de Corti embora complicada é essencialmente a mesma que a mácula e crista do vestíbulo, e parece de modo geral com outros órgãos sensoriais de origem epitelial. Aqui, também há 2 tipos de elementos histológicos. Um elemento, de uma natureza indiferente e que tem um papel suplementar compreende as células suportes, que formam o sistema de sustentação. Há, ao lado em contato com as células ciliadas, fibras nervosas cujos impulsos são transmitidos ao cérebro. Não estão presentes vasos sanguíneos e a nutrição das células do órgão de Corti se realiza por meio da endolinfa. As células de sustentação são de vários tipos. Existem as chamadas pilares interno e externo ou fibras de Corti. Elas foram nomeadas pilares pelo fato de serem retas. As tonofibrilas espessas, rígidas que elas incluem em seu arranjo é que obviamente sugerem uma função de suporte. Suas extremidades inferiores são alargadas e descansam sobre a membrana basilar. A distância entre a base do pilar externo é aproximadamente igual aos seus comprimentos. Nas duas extremidades superiores os pilares convergem e ligam-se com a superfície da papila basilar. Em secções verticais da papila o espaço livre entre os dois pilares chamado túnel interno, aparece triangular na forma, um lado do triângulo correspondendo à membrana basilar. Descansando sobre a membrana basilar estão os corpos dos dois pilares, que contêm núcleos vesiculares. Quando vistas de lado, as paredes do túnel interno não aparecem sólidas pois os pilares arranjados um ao longo do outro não formam uma linha contínua mas mais uma paliçada, com fendas livres entre as quais passam o endolinfa e as fibras nervosas. Nos topos e bases, contudo, os pilares adjacentes são contíguos. Para dentro, em direção ao lábio da lâmina espiral óssea, a papila basilar ou órgão de Corti termina com uma fileira de células falangeanas internas e as células limitantes de Held. Os dois tipos representam elementos transitórios de suporte que formam, com os pilares internos, o arcabouço onde se sustentam as células ciliadas internas. Além das células limitantes o epitélio torna-se rapidamente reduzido ao epitélio indiferenciado que limita o sulco espiral interno. Para fora dos dois pilares e do túnel interno, as células suportes são mais numerosas. Na espira basal na coclea elas estão presentes em três fileiras, na espira média em quatro e na espira apical em cinco fileiras paralelas longitudinais. Os corpos de citoplasma contendo núcleo, descansam sobre a membrana basilar, e constituem as tonofibrilas espessas, rígidas e fibrosas. Por causa da semelhança de suas extremidades superiores livres, ou cabeças, com as falanges ósseas dos dedos da. mão humana, estas células suportes têm sido chamadas células falangeadas externas, também conhecidas como células de Deiters. Aproximadamente na altura média da papila basilar as fibras das células falangeanas externas produzem pés laterais, um para cada célula. Os tipos destes processos fibrosos secundários são alargados em proeminência chamadas cabeças inferiores ou ninhos em forma de taças para o suporte das extremidades arredondadas inferiores das células ciliadas externas, cada célula falangeana suportando uma célula ciliada. Pela ligação de uma com a outra e com as extremidades superiores dos pilares, as cabeças superiores das células falangeanas formam o plano mais superficial da papila basilar. Este plano, a membrana reticular de Kolliker, não é contínuo, mas constitui uma rede em cujas malhas em forma de anel estão ajustadas os extremos superiores das células ciliadas. As extremidades inferiores e corpos das células falangeanas externas fixam-se à membrana basilar, e suas extremidades superiores, que formam a membrana reticular, tanto quanto as células ciliadas que elas suportam, estão, ambas, transversalmente e no sentido longitudinal espiral, separados por espaços intercelulares na forma de fendas estreitadas e túneis largos. O mais externo destes intersticios é chamado pilar externo ao passo que as fendas entre o túnel interno e externo são chamadas espaços de Nuel. Estes espaços são usados para a passagem das fibras nervosas e para o fluxo livre de endolinfa pelo qual todas as estruturas, pilares, células falangeanas, células ciliadas e nervos são banhadas e nutridas. Para fora, em direção à periferia do duto coclear, as células suportes, aqui chamadas células de Hensen, são ainda de uma forma colunar mesmo, embora elas não contenham estruturas como pilares (tonofibrilas como reforço). Além dos limites do órgão de Corti estas células, algumas das quais foram descritas por Claudius, são reduzidas a modestas proporções de um plano epitelial simples estreito cobrindo a face vestibular da membrana basilar até sua fixação ao ligamento espiral.
A parede lateral do duto coclear é a parede vascular. É lineada por um epitélio relativamente espesso no qual, vasos sanguíneos minúsculos penetram no ligamento espiral. Nela se localiza a estria vascular. Em suas estruturas a parede vascular parece com o plexo corioide dos ventrículos cerebrais e tem uma função semelhante; produção de endolinfa.
Células sensoriais do órgão de Coril: (Figs. 12, 13, 14, 15)
Os elementos sensoriais essenciais do órgão de Corti: as células ciliadas são sustentadas pelas estruturas já estudadas anteriormente. Estes elementos, são de dois tipos; as células ciliadas externas e as células ciliadas internas.
As células ciliadas externas têm a forma oblonga. O citoplasma contém um núcleo arredondado semelhante em forma e tamanho às das outras células epiteliais adjacentes como as células falangeanas e os pilares. São mais numerosas que as internas 12.000 na cóclea humana.
Células ciliadas internas têm a forma de um frasco, reto nas extremidades superiores e com terminações inferiores arredondadas. Os núcleos são semelhantes aos das células ciliadas externas. Estão presentes em uma fileira e são 3.500 na cóclea humana. Os extremos superiores destes dois tipos celulares são mantidos em posição pela membrana reticular.
As células ciliadas têm algumas características particulares; em primeiro lugar devemos citar os cílios de onde provém o nome. Existem várias dúzias por células, sendo mais numerosas no homem que em outros mamíferos. O comprimento dos cílios varia, mesmo na própria célula e entre diferentes células. Aumenta de 40 na espira basal a 100 na espira apical. Outra característica das células ciliadas é a presença de uma rede delicada de fibrilas argentófílas no seu interior semelhante às nervo-fibrilas das células nervosas. Estas fibrilas entretanto não tem relação direta com as fibras do nervo acústico, cujos botões terminais estão ligados do lado de fora das células ciliadas.

FIG. 12 - órgão de Corti. Vista superior.
CH - Células de Hensen
CFE - Células falangeanas externas
CCE - Células ciliadas externas
PI - Pilar interno
CCI - Células ciliadas internas
CFI - Células falangeanas internas
PEI - Plexo espiral interno
MB - Membrana basilar
PE - Pilar externo
HP - Habenula perforate
ZP - Zona pectinada
ZA - Zona arcuada
TC - Tecido conjuntivo
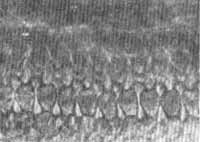
FIG. 13 - Fotomicrografia de contraste de fase do órgão de Corti pela técnica de preparações de superfície ao nível dos cílios x 1.400.
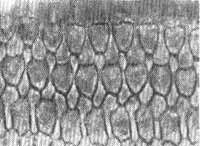
FIG. 14 - Fotomicrografia de contraste de fase do órgão de Corti ao nível dos órgãos celulares. Observam-se as 3 fileiras de células ciliadas externas. Mesma técnica anterior x 1.400.
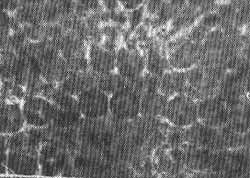
FIG. 15 - Fotomicrografia de contraste de fase do órgão de Corti. Mostrando os núcleos. Mesma técnica anterior x 1.400.
Relação dos cílios com a membrana tectorial
Alguns investigadores referem que os cílios apenas locam a membrana tectorial na sua superfície inferior. Outros afirmam que os cílios se fundem com a substância da membrana. De qualquer modo é quase certo que não terminam livres e sim que estão relacionados direta e fisicamente à membrana tectorial.
Inervação da cóclea (Fig. 16)
A secção central do nervo acústico contém neuroglia, a secção periférica tem bainha de neurilema ou células de Schwann. Os feixes deste nervo são mantidos juntos por planos de tecido conjuntivo endoneuro e perineuro que os ligam à bainha externa e epineuro.
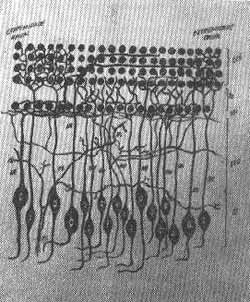
FIG. 16 - Esquema simplificado da inervação do órgão de Corti.
CCE - Células ciliadas externas
CCI - Células ciliadas internas
LEO - Lâmina espiral óssea
GE - Gânglio espiral
NE - Neurônio espiral
FS - Fibras simpáticas
ON - Orto neurônios
Gânglio coclear
As células nervosas cujas fibras formam o nervo coclear estão reunidas no osso petrosq formando o gânglio espiral. São células bipolares. De cada célula do gânglio espiral nasce um processo central e um periférico. 0 processo central torna-se, no curso do desenvolvimento, o cilindro eixo ou fibra nervosa. As fibras juntas de todas as células formam o nervo coclear. Os processos periféricos tornam-se, no desenvolvimento, extensos e lisos, assumindo a aparência de um cilindro eixo.
Suprimento nervoso do órgão de Corti
A penetração dos processos periféricos ou dentríticos das células nervosas do gânglio espiral na papila basilar é acompanhada de uma germinação ativa durante o desenvolvimento embriológico. Deste modo se estabelece uma íntima relação entre as terminações das fibras do nervo coclear e o órgão de Corti, relação esta altamente elaborada nas cócleas desenvolvidas do homem e outros vertebrados de boa audição. Esta relação é de suprema importância para a função específica da cóclea e para o receptor periférico da audição.
Os processos dentríticos referidos são dispostos em feixes que se dispõem radialmente ao eixo do modíolo, passando através de canais existentes na lâmina espiral óssea do lado vestibular. Alcançam em seguida a papila espiral basilar ou órgão de Corti no duto coclear membranoso através do lábio vestibular do limbus laminar. Quanto ao arranjo dos terminais nervosos no órgão de Corti, infelizmente, o seu conhecimento é ainda incompleto. A descrição que daremos é baseada em fatos já estabelecidos aparentemente no presente.
Das terminações nervosas da cóclea humana e outros mamíferos existem algumas diretamente relacionadas com o órgão de Corti sendo por isso considerados como fibras nervosas auditivas verdadeiras. Outras são relacionadas indiretamente com o órgão de Corti.
a) otoneurônios: são os mais numerosos. Alcançam a papila basilar em feixes dispostos radialmente sem grandes desvios. São também chamados fibras radiais ou diretas. Quando penetram na papila basilar seguem um curso reto dando poucas ramos colaterais finos que entram em contato com a superfície inferior das células ciliadas internas. Em seguida cruzam o túnel interno e espaço de Nuel e se dividem em vários ramos secundários e terciários produzindo uma arborização terminal restrita. Estas ramificações desta arborização apresentam botões terminais com os quais aderem à superfície inferior e aos lados das células ciliadas externas. Assim cada fibra periférica deste tipo de célula bipolar do gânglio espiral está relacionada a um pequeno grupo de células ciliadas externas e internas.
b) espironeurônios ou fibras espirais: estas fibras são mais espesas que as anteriores. São, entretanto, menos numerosas. A arborização terminal desta fibra fica a uma certa distância do ponto de penetração na papila basilar. O curso é radial, mais ou menos reto do gânglio espiral à papila basilar, em seguida muda abruptamente seu curso de direção para retornar longitudinalmente. Na papila basilar as terminações são encontradas em feixes por entre e abaixo das fileiras de células ciliadas externas. Do mesmo modo que as fibras já descritas, a uma certa distância do ponto de entrada da fibra, na papila basilar, eles terminam em arborização com ramos secundários e terciários. Estes ramos também apresentam botões terminais que tocam a superfície das células ciliadas externas por baixo e ao longo dos lados. Cada fibra espiral liga-se a um grupo de células ciliadas de três, quatro ou mais fileiras, sendo relacionada a poucas células ciliadas . internas através de alguns ramos colaterais vistos no ponto de entrada na papila.
c) platineurônios - fibras espirais internas, fibras difusa: são finas com arborização pequena espalhando sobre um grande segmento da cóclea, e, aparentemente, só terminam nas células ciliadas internas.
Fibras não auditivas
São simpáticas e encontradas no modíolo e na lâmina espiral óssea. Variam em espessura e por divisões repetidas produzem arborização que, juntas, formam um plexo com numerosos ramos que terminam por botões. Estas fibras terminam a certa distância da papila basilar em um tecido que contém vasos. Isto sugere que estas fibras não pertenciam a variedade "auditiva", sendo mais participantes do sistema vasomotor da cóclea.
BibliografiaEngstrom, H., Ades, H. W., Hawkins, J. E. Jr.: Cytoarquitecture of lhe organ of Corti. Acta Oto-Laryng., 1963, suppl. 188 : 92-99.
Guild, S. R.: A graphic reconstruction method for the study of the organ of Corti. Anat. Rec., 1921, 22 : 141-157.
Guild, S. R., Drowe, S. J., Bunc, H. C. C., Polvogt, L. M.: Correlations of differences in the density of innervation of the organ of Corti with differences in the acuity of hearing, including evidence as to the location in the human cochlea of the receptor for certain tones. Acta OtoLaryng., 1931, 15 : 269-308.
Held, H.: Die Cochlea der Sãuger und der Vógel, ihre Entwicklung und ihr Bau. In: Bethe, A., von Bergmann, G., Embden, G., Ellinger, A. (Ed)., Hanbuch der Normalen und pathologischen Physiologie, vol. II, Springer, Berlin, 1926 : 467-534.
Oliveira, J. A.: Anatomo fisiologia animal comparada. Editora Universitária. Ribeirão Préto, 1971, 414 pp.
Oliveira, J. A., Marseillan, R. F.: A técnica de preparação de superfície e sua aplicação no estudo da citoarquitetura normal e patológica do órgão de Corti. Comunicação ao XVII Congresso Brasileiro de Otorrinolaringologia, Brasília, 1968.
Oliveira, J. A. e Marseillan, R. F.: Estudo das lesões cocleares pela técnica de preparações de superfície na cobaia. Revista Brasileira de Otorrinolaringologia, 1971, 37 : 1.
Retzius, G.: Gehõrorgan der Wirbeltiere. II. Das Gehororgan der Reptillien, der Vogel und der Saugetiere. Samson & Wallen, Stockolm, 1884.
Schucknecht, H. F.: Techniques for study of cochlear function and pathology in experimental animais. Arch. Otolaryng., 1953, 58 : 377-397.
Vinnikov, Y. A., Titova, L. K.: The organ of Corti. Its Histophysiology and Histochemistry. Consultants Bureaus, New York, 1964, 253 pp.
* Trabalho efetuado com auxílio da Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP).
** Professor Assistente Doutor contratado do Departamento de Oftalmologia e Otorrinolaringologia da Faculdade de Medicina de Ribeirão Prêto da U.S.P. Professor Assistente Doutor da Disciplina de Anatomia e Fisiologia Humana da Faculdade de Filosofia, Ciências e Letras de Ribeirão Prêto.
