

Ano: 1974 Vol. 40 Ed. 1 - Janeiro - Abril - (10º)
Seção: Artigos Originais
Páginas: 64 a 82
FISIOPATOLOGIA DO SISTEMA VESTIBULAR (*)
Autor(es): José Antonio A. Oliveira (**)
![]()
Consideramos o objetivo do presente trabalho o conhecimento dos fenômenos fisiológicos básicos para um perfeito entendimento da patologia do sistema vestibular de modo que o otorrinolaringologista possa a luz destes fenômenos compreender as alterações da fisiologia normal do sistema através dos sintomas e sinais patológicos existentes. Não nos deteremos em processos ainda não esclarecidos ou teorias, pois o assunto é bastante extenso e com issc estaríamos fugindo do objetivo.
FISIOPATOLOGIA DO SISTEMA VESTIBULAR
Consideremos o sistema vestibular com suas vias e suas conexões funcionando como um arco reflexo.
Como se conhece de neurofisiologia o arco reflexo é formado pela seqüência: (A) Estímulo; (B) Receptor; (C) Vias nervosas aferentes; (D) Centro nervoso; (E) Vias eferentes; (F) Efetor; (G) Resposta.
A - Estimulo
O labirinto posterior constituido pelo sáculo, utrículo e canais semicirculares é responsável fundamentalmente pela recepção de estímulos que nos informam a posição da cabeça no espaço e o sentido de rotação da mesma.
As experiências de Huizinga (1955) demonstraram que principalmente o utrículo tem sua atividade de repouso modificada com a mudança de posição da cabeça. Deste modo podemos dizer que um dos estímulos para os receptores labirínticos é a aceleração linear e centrífuga, como é o caso da gravidade.
Desde as experiências de Cum-Brown, e Brewer, (1874-1891) evidenciou-se a teoria hidrodinâmica que relacionava o funcionamento dos canais semi-circulares com o deslocamento da cabeça em um dos três planos do espaço. Estes movimentos de rotação, devido a aceleração angular da cabeça provocariam movimentos da endolinfa no interior dos canais. Esta teoria foi em 1892 demonstrada por Ewald. Atualmente as pesquisas para estudos dos limiares de excitabilidade dos receptores dos canais semicirculares têm sido usadas cadeiras rotatórias com acelerações mínimas de até 0,001.º/S2 (Clark, 1970).
B - Receptor
1 - Estrutura do sistema vestibular (Fig. 1)
Labirinto posterior ósseo - está localizado no osso temporal na parte petrosa.
É constituído por cavidades afuniladas. Existem os três canais semi-circulares cada um com duas extremidades que se abrem numa câmara ovóide, o vestíbulo.
Labirinto posterior membranoso - É formado por três canais semi-circulares alojados nos canais ósseos membranosos correspondentes e duas vesículas membranosas, o sáculo e o utrículo que se localizam no vestíbulo ósseo.
Canais semi-circulares - Situam-se em planos que formam aproximadamente entre si ângulos retos. São denominados canal externo horizontal ou lateral - canal anterior (vertical) e canal posterior (vertical). Todos os, canais apresentam numa de suas extremidades uma ampola onde se localiza o órgão receptor. Todas as estruturas labirínticas membranosas estão rodeadas de perilinfa e contêm endolinfa no seu interior.
FIG. 1 - Estrutura do Labirinto Membranoso.
O canal horizontal apresenta sua convexidade para fora e para trás. A cabeça estando em posição ereta, este canal forma com o plano horizontal um ângulo de 30° com abertura anterior. Deste modo o plano do canal é desviado para cima do plano horizontal. Por este motivo por ocasião do teste calórico deve-se corrigir a cabeça do paciente elevando-a a uma posição que corresponde a uma angulação de 30°, estando o paciente em decúbito dorsal.
Os canais verticais (anterior e posterior) fazem ambos um ângulo de 45° com os planos frontal e sagital do crânio, isto é, seus planos atravessam diagonalmente os planos frontal e sagital. Deste modo o canal anterior de um ouvido está no mesmo plano que o canal posterior do outro ouvido, enquanto o canal posterior ou anterior de um ouvido forma um ãngulo reto com seus companheiros do lado oposto.
Os canais laterais de ambos os lados ficam no mesmo plano.
Sáculo e utrículo - Estão na cisterna perilinfática (vestíbulo).
O sáculo está localizado no recesso esférico na parede medial do vestíbulo. Sua parte superior se projeta para o utrículo e se liga ao recesso do utrículo sem haver comunicações entre as duas cavidades.
O sáculo está ligado ao canal coclear pelo duto "reuniens", um fino canal que deixa a parte póstero-inferior do sáculo, terminando na parte basal do canal coclear.
Do sáculo sai ainda um canal (canal sacular) em uma direção súpero-posterior que recebe um canal do utrículo (canal utricular) formando os dois o canal endolinfático que termina na cavidade craneana na superfície posterior da parte petrosa do osso temporal entre os dois planos da duramater.
O utrículo é um ducto de forma irregular com uma secção oval. Localiza-se no recesso elíptico na parede medial do vestíbulo acima do sáculo. Tem a forma aproximada de um rim, sendo a parte anterior um pouco mais larga que a posterior. Tem seis aberturas; uma para cada das três ampolas dos canais semi-circulares, uma para o duto utricular, uma para a extremidade não ampolar do canal semi-circular horizontal e uma para a "crus crommuni" abertura única para os dois canais verticais.
Constituição e localização dos receptores
Ampola membranosa - A parte basal da ampola fica em contacto com o osso, por ali penetra o tecido conjuntivo com vasos e fibras nervosas para a luz da ampola. Este tecido conjuntivo tem a forma de uma crista, a crista ampolar que é coberta com epitélio sensorial. Sobre o epitélio sensorial está uma substância gelatinosa, a cúpula que se extende ao teto da ampola e as suas paredes laterais. É formada por uma rede de fibras com diâmetro de 100A.º. É constituída de sulfomucopolisacarídeos suportados em uma matriz proteica (Vilstrup, 1950, Plotz and Perlman, 1955). Ela apresenta canais onde se introduzem os cílios das células sensoriais. (Steinhausen, 1933).
Crista ampolar - É formada por um grupo de células ciliadas sensitivas entre as quais se dispõem células de sustentação. Sobre a crista encontramos uma cúpula gelatinosa.
Estrutura do epitélio sensorial - Nos últimos anos pesquisas com técnicas microscópicas modernas, como a microscopia eletrônica a microscopia com contraste de fases e as observações tridimensionais com o "scanning electron microscope" permitiram o conhecimento da estrutura celular do epitélio sensorial dos receptores vestibulares.
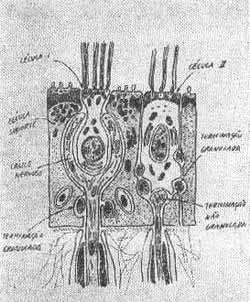
Wersali (1956) descreveu dois tipos de células sensoriais na crista ampolar da cobaia.
A célula sensorial Tipo I tem a forma de um vaso e é rodeada por um cálice nervoso vindo de fibras nervosas espessas do nervo vestibular. Cada fibra destas inerva de 2 a 5 células sensoriais do Tipo I.
A célula sensorial Tipo II é uma célula cilíndrica inervada por terminações nervosas pequenas vindo de fibras finas terminando na base da célula. (Fig. 2).
FIG. 2 - Células sensoriais dos receptores vestibulares.
Cílios - (Fig. 3) - Emergem da superfície celular e tomam contacto com
estruturas receptoras auxiliares que são:
a) na ampola dos canais semi-circulares - a cúpula;
b) no sáculo e utrículo - a membrana otolítica (sacular e utricular).
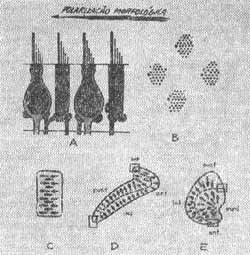
MORFOLOGIA DOS PELOS SENSORIAIS - (Wersali, 1956)
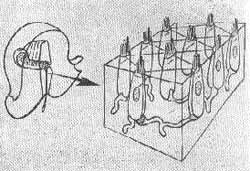
Estereocilios - Estão agrupados regularmente de 50 a 100 por célula na cobaia, seus comprimentos diferem sendo o mais longo localizado junto ao kinocilio. Esta variação de comprimento é gradual diminuindo os cílios a medida que se afastam do kinocilio.
FIG. 3 - Disposição dos estereocilios e kinocilios na célula receptora vestibular e do órgão de Corti.
É o estereocilio formado por uma membrana periférica constituída por três planos circundando um centro fibrilar protoplasmático que continua em sua base com um filamento axial que atravessa a cutícula da célula.
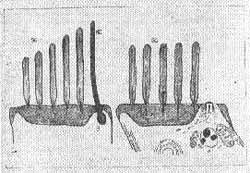
Kinocilio - É constituído de um conjunto de filamentos que nascem num corpo basal abaixo da superfície celular. Existe um para cada célula. Existe um par central de filamentos rodeado de nove filamentos tubulares duplos periféricos ligados no corpo basal da célula. O kinocílio é limitado por uma membrana que é contínua com a membrana plasmática da célula. O diâmetro total do kinocílio é de 0,2 a 0,4 micras e varia com o órgão terminal e com o animal. Tem 18 a 20 micras de comprimento na cobaia. Orientação dos cílios - Lowestein e Wersall (1959) trabalhando com peixes cartilaginosos (raia) mostraram que os feixes de pelos sensoriais têm uma distribuição particular. Sempre o kinocílio se localiza na periferia.
Na ampola do canal horizontal o kinocilio está dirigido em direção ao utrículo, nas ampolas dos canais verticais está em posição contrária, oposta ao utrículo (Fig. 4). Engstrom e colaboradores (1962) observaram uma orientação uniforme nos feixes de cílios na mácula utricular.
FIG. 4 - Receptor ampolar - indicação da polarização morfológica das células sensitivas.
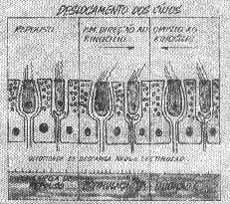
Utrículo - Encontramos a mácula utricular coberta pela membrana otolitica. Esta é carregada de otoconias incrustadas.
Os otoconios ou otolitos são cristais de calcita ou aragonita (Carlström e col. 1953). A mácula utricular tem a mesma constituição da crista da ampola dos canais semicirculares. Dispõe-se no plano horizontal quando o indivíduo está com a cabeça na posição ereta entretanto a parte anterior fica discretamente elevada.
Polarização morfológica como na crista, a célula sensorial é morfologicamente polarizada e a direção da polarização é determinada pela posição do kinocílio em relação aos estereocílios da mesma célula. Mesmo que a polarização morfológica da célula possa variar em alguma extensão, isto não altera a orientação principal na área. A mácula do utrículo é dividida em duas áreas: a parte externa e interna por uma linha curva imaginária. As células sensoriais em cada uma das duas metades são polarizadas em direção a esta linha. Os kinocilios estão voltados para ela. (Fig. 5). Sáculo - A mácula tem constituição semelhante a do utrículo. Na posição anatômica normal da cabeça, no homem na posição ereta ela é aproximadamente vertical e está situada obliquamente com um ângulo de 30.º do plano vertical. Deste modo os otolitos ficam laterais aos cílios.
FIG. 5 - Mácula utricular coberta pela membrana otolitica. Observa-se a polarização das células sensoriais com os kinocilios dirigidos para a linha média.
FIG. 6 - Mácula sacular coberta pela membrana otolitica. Observa-se a polarização das células sensoriais com os kinocilios dirigidos distalmente à linha média.
A mácula do sáculo também pode ser dividida em duas áreas: a parte externa e a parte interna por uma linha curva imaginária. A polarização aqui ocorre no sentido de se afastar da linha divisória, orientando-se deste moda para a periferia. Os kinocílios estão do lado oposto da linha divisória (Fig. 6).
MECANISMO DE ESTIMULAÇÃO
O indivíduo estando com a cabeça numa certa posição no espaço tem as células sensitivas das suas máculas saculares ou utriculares, estimuladas pelo peso da membrana otolítica (aceleração linear - gravidade).
Do mesmo modo quando se gira a cabeça esta aceleração linear provoca deslocamento de endolinfa dentro de um ou mais canais semicirculares e conseqüentemente havendo deslocamento da cúpula e conseqüentemente dos cílios.
Deste modo as forças que atuam sobre o labirinto são transferidas às terminações ciliadas das células sensoriais pela ação da cúpula gelatinosa ou membrana otolítica.
Deve-se salientar que aceleração linear e força gravitacional têm pouco efeito na cúpula. Tais forças são efetivas para os otolitos que têm maior peso. Deslo- camento dos otolitos causados por mudanças em posição afeta a atividade das células sensoriais até que o equilíbrio gravitacional normal seja restabelecido.
POLARIZAÇÃO MORFOLÓGICA
(Figs. 7 e 8)
Registrando potenciais de ação de fibras unitárias da crista (Lowestein and Sand, 1940; Adrian, 1943) e da mácula (Lowestein and Roberts, 1950), mostraram que mesmo quando a cabeça não se movia, impulsos eram conduzidos numa direção central e verificaram que esta atividade expontânea estava presente na maioria das fibras nervosas. Os deslocamentos da cúpula ou membranas otolíticas alteravam esta atividade nas fibras nervosas. Quando o deslocamento da cúpula na crista lateral era na direção utriculópeta (em direção ao utrículo) havia despolarização do epitélio sensorial (Trincker, 1957) e aumento de impulsos nas fibras nervosas (Lowestein and Sand, 1940). Deslocamento utriculófugo da cúpula produz hiperpolarização do epitélio sensorial e decréscimo da atividade das fibras nervosas. Condição invertida foi encontrada nos dutos unicirculares verticais. Em registros da mácula do utrículo (Trincker, 1957), deslocamento da membrana estatoconial (otolítica) em uma direção produz hiperpolarização, na direção oposta despolarização.
Estes achados vieram demonstrar que o epitélio sensorial vestibular é funcionalmente polarizado. Isto foi correlacionado com a morfologia das células sensoriais e disposição no órgão receptor quando Lowestein and Wersall (1959) estabeleceram que os feixes de cílios na crista lateral eram orientados em direção ao utrículo e os da crista anterior em direção oposta. A estimulação nos pelos sensoriais em uma direção do estereocílio para o kinocílio provoca despolarização da célula sensorial e um aumento na freqüência de impulsos nas fibras nervosas (Flock and Wersall, 1962; Dyjkgrasf, 1963). Além disso Flock, (1965) referiu que o efeito de qualquer força era proporcional ao tamanho da componente na direção de polarização da célula sensorial.
FIG. 7 - A - Disposição dos cílios. B - Estereocíiios e kinocílios. C - Polarização dos cílios na crista ampolar. D - Polarização dos cílios na mácula sacular. E - Polarização dos cílios na mácula utricular.
FIG. 8 - Mecanismo eletro fisiológico de estimulação.
A influência nas células sensoriais de um componente agindo tangencialmente à superfície do epitélio (Shearing force), resulta na produção de um potencial gerador nas células sensoriais. Este potencial é proporcional ao grau de deslocamento da cúpula (Flock, 1965). O mecanismo de transferência de energia às células sensoriais não é ainda conhecido.
O esterocilio ao microscópio eletrônico dá impressão de um fio rígido, acredita-se que estes cílios atuariam como microalavancas transferindo energia à superfície cuticular e às organelas na parte superior da célula (Engström et al. 1962; Lowestein and Osborne, 1964).
Flock, 1965; Wersall, 1967; Ades and Engström 1965 consideram o kinocílio e corpo basal respectivamente como as estruturas essenciais excitáveis na estimulação da célula sensorial e dirige a atenção a presença destas estruturas em outros órgãos dos sentidos e no sistema nervoso central. Sugere também que o kinocílio nas células sensoriais vestibulares pode atuar como uma estrutura móvel respondendo a uma deformação passiva com a iniciação de uma modificação elétrica (Lowestein and Wersall, 1959) (Lowestein, 1966). Deste modo um deslocamento do kinocílio perpendicular a uma linha através seus dois filamentos centrais em direção ao corpo basal, poderia estimular as células sensoriais.
Apesar destas teorias na realidade não se conhece como a energia é transferida as células sensoriais vestibulares ou como os mecanismos receptores no ouvido interno convertem energia mecânica em impulsos nervosos. Nem é conhecido se é a posição excêntrica do kinocílio e corpo basal ou o comprimento gradual decrescente do esterocílio que é responsável pela sensitividade direcional das células. Estudos posteriores ao nível ultraestrutural poderão conduzir a um melhor entendimento deste problema.
Qualquer deslocamento da membrana otolítica tangencialmente a superfície do epitélio sensorial sempre dará um estímulo excitatório a alguns grupos de células, um inibitório em outro. Parece claro também que o processo de impulsos nas fibras nervosas aferentes da mácula do utrículo deva ser muito complicado.
Com a cabeça na posição anatômica normal, a mácula do sáculo tem uma posição quase vertical e as células sensoriais são orientadas em uma direção póstero-superior e antero-inferior: deste modo com a cabeça na posição anatômica normal, uma freqüência relativamente alta de impulsos constantes nas fibras nervosas da parte externa onde a direção da polarização coincide com a direção da gravidade. Ao mesmo tempo espera-se inibição da atividade expontânea nas fibras da parte interna.
INTERPRETAÇÃO DA ESTIMULAÇÃO DAS CÉLULAS RECEPTORAS BASEADA NA POLARIZAÇÃO DAS CÉLULAS
Wersall (1961), Flock e Wersall (1962) correlacionando tabelas eletrofisiológicas com seus achados da orientação das células sensoriais polarizadas (orientação do kinocílio em relação aos estereocílios) demonstraram uma base morfológica de sensitividade direcional das células receptoras.
Lowestein e Wersall (1959) propuseram que a resposta das células ciliadas é determinada pela direção que o estímulo chega ao feixe de cílios. Deste modo a célula tem uma sensitividade direcional devido a orientação dos cílios, tal que um deslocamento da cúpula no sentido do estereocílio para o kinocílio é acompanhado de um aumento da velocidade de descarga na fibra nervosa aferente; na direção oposta o deslocamento cupular produz efeitos contrários.
C - Vias nervosas aferentes
As terminações nervosas Tipo I já descritas com as células sensoriais tem poucas vesículas grandes. As do tipo II contêm uma acumulação densa de vesículas de modo semelhante as encontradas em terminações pré-sinápticas no sistema nervoso central. Estudos recentes têm demonstrado a existência de estruturas sinópticas típicas na junção entre terminação nervosa e célula sensorial.
SIGNIFICADO DAS DIFERENTES TERMINAÇÕES NERVOSAS
Os cálices nervosos das células ciliadas Tipo I e as terminações nervosas não granuladas de células ciliadas Tipo II são consideradas terminais sensoriais aferentes.
Associado a este tipo de terminação nervosa é visto uma faixa sinóptica de estrutura típica.
Engström (1958) discute a existência de sistema bem desenvolvido de fibras aferentes e eferentes na cóclea como nas áreas vestibulares sensoriais. Comparou a estrutura das terminações nervosas vesiculosas com terminações pré-sinópticas no sistema nervoso central e sugeriu que todas terminações nervosas vesiculosas eram os terminais de fibras eferentes. Estudos por Dohlman and Anggärd (1958) demonstraram a existência de ácetilcolinesterase específica no epitélio sensorial vestibular, um fato que foi considerado um sinal de atividade eferente. Hilding and Wersall (1966) localizaram a acetilcolinesterase no ouvido interno nas fibras nervosas granuladas. Nenhuma esterase foi encontrada nos cálices.
Rasmussen (1946) mostrou a existência de fibras nervosas eferentes dos núcleos vestibulares a todo epitélio sensorial vestibular.
lurato (1962), Kimura and Wersall (1962) mostraram que cortando o trato olivo-coclear cruzado causa a degeneração das terminações nervosas granuladas nas células ciliadas externas do órgão de corti. O significado fisiológico dos dois diferentes tipos de células sensoriais no sistema vestibular não é conhecido.
Parece que as células do Tipo I inervadas por cálices nervosos grandes de fibras nervosas grandes (grossas) podem ser olhadas como mais diferenciadas, altamente sensitiva suprindo a resposta local dentro de uma pequena área do epitélio. As células Tipo II menos especializadas e difusamente dispostas podem provocar informação de caráter geral.
nvestigações clínicas sugerem que atividade eferente afeta a resposta das células receptoras do sistema vestibular. Atividade eferente aumentada parece ser a causa de inibição da descarga aferente espontânea decréscimo da excitabìlidade e menor potencial de ação. Atividade eferente reduzida, de outro lado, produz um aumento na freqüência de descarga aferente e aumento na excítabilidade.
As fibras nervosas aferentes que provêm das cristas ampolares ou das máculas otolíticas atingem a cavidade craneana pelo meato acústico interno onde se encontram os corpos celulares bipolares formando o gânglio vestibular ou de Scarpa. Estas fibras vão formar a porção vestibular do oitavo nervo craneano que unindo-se a porção coclear se dirigem aos núcleos bulbares seguindo o mesmo trajeto. Este neurônio representa o primeiro neurônio vestibular. Do meato auditivo interno seguem as fibras até os núcleos vestibulares localizados no bulbo próximo a linha média em relação com o assoalho do IV ventrículo inferiormente.
D - Centros nervosos
São os núcleos vestibulares bulbares de Deiters, Schwalbe, Bechterev, Roller, Brodal. Nestes núcleos encontramos as células do segundo neurônio. O segundo neurônio vestibular é um neurônio de associação com os núcleos oculomotores, medulares neurovegetativos e constitue o "cierre" do arco reflexo vestíbulos efetor, muscular ou neurovegetativo.
E - Vias eferentes efetoras Conexões vestibulares, conexões musculares.
Conexões oculomotores. (Vias vestíbulo-oculomotores).
A projeção do estímulo vestibular aos núcleos oculomotores produz a coordenação dos olhos com a posição da cabeça e corpo no espaço. O aumento da freqüência de descarga de impulsos originadas por exemplo nas células receptoras da crista ampolar do canal horizontal é transmitido aos núcleos oculomotores contralaterais pois as fibras que saem dos núcleos vestibulares cruzam a linha média antes de alcançar o núcleo do nervo craneano motor ocular comum (Ill) e do nervo abducente (VI). O primeiro inerva o músculo reto medial do lado oposto ao núcleo e o segundo o reto lateral do mesmo lado.
Conexões medulares (vias vestíbulo-espinais)
Estas conexões se realizam em grande parte através do núcleo de Deiters que recebe também fibras de cerebelo de modo que o sistema vestibular e o cerebelo juntos possam regular o tono muscular do pescoço, troncos e extremidades de ambos os lados. O aumento da freqüência de descarga por exemplo nas células receptoras da crista do canal horizontal determina a transmissão do impulso (potencial de ação) até os núcleos dos nervos espinais do lado oposto principalmente o que determina o aumento do tono em toda musculatura contralateral ao lado estimulado, musculatura esta do pescoço, tronco, extremidades.
Conexões cerebelosas
O cerebelo recebe o estímulo proprioreceptivo vestibular, tanto do primeiro neurônio como dos núcleos de Betcherew e Deiters. Estes impulsos provenientes do sistema vestibular se associam aos impulsos proprioreceptivos da sensibilidade profunda e estímulos estero e interoceptivos do organismo. Deste modo pela via cerebelo-vestíbulo-núcleo de Deiters espinal, mantém o tono muscular adequado para conservar o equilíbrio e os movimentos.
Conexões neurovegetativos - São múltiplas.
a) com o núcleo vegetativo hipotalãmico
b) com a substância reticular bulbar e mesencefálica
c) com o pneumogástrico (vago)
Conexões corticais
O sistema vestibular é um órgão sensorial e para tanto deve ter um componente consciente. A mais importante via parece ser a que envia ao cerebelo, região talâmica e cortical o impulso proveniente de estímulos proprioceptivos vestibulares e de sensibilidade profunda .
Estes estímulos de sensibilidade profunda devidos a contração muscular, estiramento de tendões e posição das articulações informam ao cortei as posições dos segmentos do corpo no espaço. A via vestibular informa a posição da cabeça estática ou dinâmica no espaço devido estímulos provenientes do sistema vestibular. O conjunto destes estímulos dá a sensação de equilíbrio.
F - Efetores
Corresponderiam aos músculos do olho, a musculatura esquelética estriada de todos os segmentos do corpo e aos núcleos do nervo vago.
G - Ação dos efetores - Resposta reflexa
Músculos do olho - Nistagmo
O aumento do tono dos músculos do olho, reto lateral do lado oposto ao estimulado e reto medial do olho do lado estimulado provoca, um desequilíbrio de tonos em relação aos músculos antagonistas e os dois olhos são desviados conjugadamente para o lado oposto ao estímulo.
Este desvio de origem vestibular corresponde a fase lenta do nistagmo e corresponde a um movimento compensatório dos olhos para reter o campo do último olhar oposto ao movimento da cabeça pois como vimos se a cabeça gira para a direita horizontalmente por exemplo este desvio é para a esquerda. Este é um mecanismo protetor retendo o sistema nervoso a orientação do meio ambiente. A mesma ação do exemplo citado pode ser obtida estimulando-se com água quente o canal horizontal direito de um paciente em decúbito dorsal com a cabeça elevada de 30° em relação ao plano horizontal.
A volta do olho em sentido contrário constitue a fase rápida do nistagmo. A direção do nistagmo é dada por este componente rápido.
Ewald (1892) foi quem determinou ao labirinto o nistagmo.
Buys (1924) e Dohlman (1925) verificaram aplicando estímulos rotatórios e calóricos respectivamente que a velocidade dos olhos durante o componente lenta é proporcional ao estímulo aplicado no sistema vestibular.
Quando o componente rápido do nistagmo McCabe (1965) conclui que para seu aparecimento a substância reticular é indispensável.
Músculos espinais
O aumento do tonos dos músculos dos segmentos do corpo do lado oposto ao estímulo, provoca desequilíbrio em relação ao tonos dos músculos do lado oposto, isto pode determinar a queda do indivíduo para o lado contrário ao estimulado.
Núcleos vagais
A freqüência de descarga dos impulsos nervosos provenientes dos receptores vestibulares podem determinar devido aos núcleos vagais, reações neurovegetativas como náusea, vômito, queda de pressão, sudorese, diarréia.
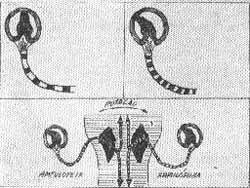
A TEORIA DO EQUILíBRIO
(Fig. 9)
Em condições normais a estimulação (rotação) provoca uma modificação da descarga de impulsos de repouso dos dois lados. De um lado o potencial de repouso é modulado num sentido e do outro em sentido contrário. A diferença de modulação é a mesma dos dois lados em sentidos contrários. Deste modo os dois lados do cérebro são informados de um modo igual mas oposto. Esta polaridade é importante para se compreender o sistema.
Esta polaridade normalmente é recebida pelas estruturas com as quais o sistema vestibular mantém conexões.
a) o cortex interpreta a modificação como um movimento na direção e velocidade específica.
b) os núcleos dos músculos oculares movem os olhos compensatoriamente para reter o campo do último olhar, oposto ao movimento da cabeça, como um mecanismo protetor retendo a orientação do meio ambiente.
c) as células do corno anterior da medula espinal ajusta o tonos dos músculos do tronco e membros.
d) o cerebelo ajusta o tono para a nova situação.
Estes processos são em parte instintivos e parcialmente aprendidos.
O cortex aprende relacionar o grau de inclinação da cúpula em resposta a movimentos repentinos da cabeça, com o grau específico de alteração do meio ambiente por equiparação; os olhos informam que o corpo moveu, os proprioceptores também, modalidades tácteis informam ao cérebro quando o organismo toca o solo. Durante anos o cérebro aprende a combinar as diferentes modalidades com o sistema vestibular originando-se então a sensação vestibular. Sendo um computador perfeito, na infância já integrou estas sensações.
FIG. 9 - Mecanismo de estimulação normal dos receptores vestibulares.
Podemos dizer que a finalidade dos dois sistemas vestibulares é a de informar o organismo de movimentos e posição da cabeça para o ajuste do corpo a estas novas posições.
Comparando este esquema de funcionamento e a estrutura do sistema vestibular podemos interpretar à luz destes conhecimentos os sintomas provocados por doenças vestibulares, sintomas estes narrados pelo paciente e os sinais que poderão ser detectados pelos testes labirínticos e eletronistagmografia.
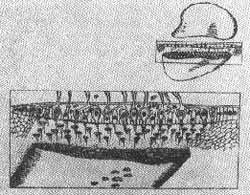
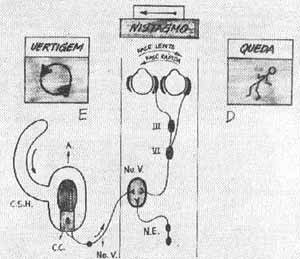
CRISE LABIRINTICA
(Fig. 10)
Quando há um distúrbio num labirinto, esquerdo por exemplo, resultando em diminuição repentina da freqüência de descarga ele é incapaz de contrabalançar o lado normal com uma descarga oposta e igual.
O cérebro capaz de interpretar a luz de respostas aprendidas passadas distribue uma falsa corrente de impulsos a seus efetores. Deste modo surge a vertigem; o cortex é informado que o organismo está em um estado constante de movimento.
Os olhos procuram repetitivamente reter o campo do último olhar (nistagmo) os músculos dos membros preparam-se para encontrar uma situação que é irreal (desequilíbrio e queda e o intestino pára e começa a peristalse reversa (náusea e vômito).
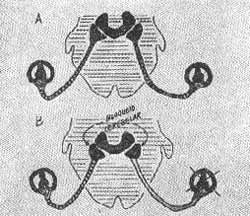
FIG. 10 - Esquema das vias e conexões dos receptores ampolares, mostrando alterações provocadas por estimulação esquerda ampolar (canal horizontal. C.S.H. Semi-circular horizontal. C.C. Célula ciliada. Nu.V. Núcleos vestibulares. N.E. Núcleos espinais.
O lado envolvido (doente, hipofuncionante como na doença de Menière, não é capaz de comunicar ao cérebro sua informação de repouso. Deste modo os dois sistemas vestibulares estão descarregando em repouso em uma intensidade desigual o que tem para o cérebro um significado específico. As sequelas deste desequilíbrio são manifestações de uma hiperfunção relativa do lado intacto; então como resultado aparecem reflexos vestibulares prolongados e incontrolados.
Cortex - interpreta o desequilíbrio como condição de movimento constante; esta é a definição de vertigem. Esta sensação pode ser de inclinação, rolamento, elevação mas sempre de natureza rotacional. Este é um sintoma comum que relata o paciente vestibular. As análises de suas características poderão trazer informações preciosas para o diagnóstico do distúrbio labiríntico.
MÚSCULOS OCULARES
O mesmo desequilíbrio de descargas chegando aos músculos oculares e a formação reticular provocam o nistagmo. Os músculos do olho desviam os olhos na direção do último olhar para reter a orientação, é a componente lenta do nistagmo. Os olhos não podem contudo continuar seguindo a pista indefi- nidamente em uma única direção por causa de suas limitações anatômicas, assim depois de um desvio específico devido os motoneurônios, neurônios inibidores na formação reticular desfazem o fluxo vindo do núcleo vestibular e ao mesmo tempo neurônios reticulares ativadores dirigem os núcleos dos músculos oculares para voltar os globos oculares ao ponto do olhar em que a componente lenta começou o desvio. Esta segunda fase do desvio do olho é muito mais rápida porque é uma fase compensatória daí o aparecimento do componente rápido do nistagmo. O neurônio reticular ativador entra em período refratário e o influxo do órgão terminal do núcleo vestibular reassume seu efeito sobre os tratos dos músculos do olho, os globos oculares são dirigidos outra vez para reter o campo do último olhar. Esta tentativa repetitiva para reter o último campe do olhar por um movimento conjugado dos olhos e um reflexo rápido de volta dos globos oculares através da linha média em compensação é o nistagmo vestibular.
O nistagmo tem um valor muito grande no diagnóstico de doenças labirínticas. A sua análise, quando provocado por provas rotatórias ou térmicas principalmente com registro eletronistagmográfico traz informações objetivas que permitem o diagnóstico da doença labiríntica.
Temos atualmente utilizado a técnica de registro eletronistagmográfico em cobaias para estudo da função vestibular, utilizando cobaias com eletrodios implantados nos cantos externos dos olhos e ligados a um conector fixo a cabeça com acrílico. Este conector fixo pode ser ligado a um amplificador e um polígrafo que registrará o nistagmo provocado por provas térmicas.
Aplicando drogas no ouvido médio de cobaias preparadas deste modo, como o salicilato de sódio (Oliveira, 1971) temos abolido a função vestibular do lado da administração ocorrendo reações reflexas idênticas as descritas atrás, ou seja uma crise semelhante à que ocorre na doença de Menière.
Deste modo podemos estudar as características eletronistagmográficas do distúrbio labiríntico experimentalmente. Também a aplicação de antibióticos vestibulotóxicos tem sido estudada por administração sistêmica crônica havendo abolição da função vestibular em tempos variáveis. Pode-se assim observar o desaparecimento do nistagmo no registro eletronistagmográfico. Os antibióticos mais tóxicos ao sistema vestibular em cobaias têm sido a gentamicina, estreptomicina. O que se confirma com estudos em preparações histológicas de superfície do epitélio vestibular (Oliveira, 1973).
MEDULA ESPINAL
A informação de desequilíbrio é transmitida do núcleo vestibular para a medula espinal, células do corno anterior, instruindo os músculos posturais e locomotores para se preparar para uma situação falsa. Deste modo ocorre a queda para o lado hipofuncionante pois o tono dos músculos de todos os segmentos do corpo deste lado está aumentado o que não é compensado pelo lado oposto.
O sentido da queda é clinicamente importante para a análise do sistema vestibular lesado.
NÚCLEO DORSAL DO X
O desequilíbrio de impulsos também chega ao núcleo dorsal do vago. No início este núcleo só efetua a cessação da peristalse. Se o equilíbrio é máximo e contínuo contudo ocorre peristalse reversa com náusea e vômito.
CEREBELO
A resposta do cerebelo ao desequilíbrio está começando a ser entendida. Em uma questão de minutos o cerebelo impõe corte virtual da atividade elétrica do núcleo vestibular em virtude de sua profunda influência inibitória da atividade vestibular.
O cerebelo não elimina o desequilíbrio por seu corte, porque nem toda informação dos órgãos terminais se distribuem ao cérebro através dos núcleos (medial principalmente); algumas fibras do órgão terminal distribuem impulsos a numerosas outras partes do tronco cerebral e cerebelo sem sinapse vestibular nuclear.
O corte nuclear não elimina então o problema, mas serve para diminuir o desequilíbrio (Fig. 11).
FISIOLOGIA DA COMPENSAÇÃO
O estado de desequilíbrio não poderia durar muito tempo, o organismo eventualmente morreria através da desidratação e desequilíbrio fluído e ele- trolítico. Deste modo o organismo tenta equilibrar a situação. A restauração do equilíbrio entre os dois centros inicia o controle dos reflexos patológicos.
Esta restauração pode ocorrer por supressão central do lado intacto por invocação de tratos inibitórios no sistema nervoso central.
FIG. 11 - Fisiologia do bloqueio cerebelar.
Também pode haver geração de uma nova atividade elétrica no sistema com descarga menor para equilibrar o lado normal, hipercinético em relação ao lado oposto pela condição patológica. Após dias ou semanas da lesão do órgão terminal esta nova atividade de repouso do núcleo denervado determina o desaparecimento dos sintomas e o corte cerebelar vai desaparecendo vagarosamente. Não se sabe o que estimula a geração de nova atividade de repouso no núcleo denervado mas sabemos que é necessário o desequilíbrio provocando uma espécie de vácuo que estimula a necessidade do seu aparecimento.
Além do mais o organismo tenta fazer o órgão doente voltar a normalidade.
Esta restauração torna a situação tolerável para o organismo desde que não haja estimulação do lado bom o que acentua o desequilíbrio. Daí durante a crise o paciente se mantém bem sem movimento da cabeça.
Isto acontece nas várias doenças vestibulares como por exemplo na Doença de Menière, labirintite supurativa etc.
A rapidez da recuperação dependerá da idade do paciente, eficiência do bloqueio neuronal, eficiência do suprimento vascular do sistema nervoso central. Este mecanismo de compensação tem uma implicação clínica muito importante; se os sintomas persistem continuamente por mais de 2 ou 3 semanas a causa não é vestibular. Este fato associado a crise significante sem nistagmo labiríntico expontâneo nos indicam que o distúrbio não é vestibular.
CONCLUSÃO
Temos condições conhecendo estes processos fisiológicos básicos, de interpretar os sistemas que o paciente apresenta quando com patologia vestibular. Sintomas como vertigem, desequilíbrío, náuseas e vômitos.
Os sinais apresentados pelo paciente podem ser analisados e interpretados por provas objetivas. É o caso do desequilíbrio que pode ser analisado por testes simples de equilíbrio estático e dinâmico e o nistagmo que pode ser registrado eletricamente utilizando-se a eletronistagmografia, nstagmo este provocado por provas calóricas e rotatórias, cujas técnica fogem ao objetivo deste trabalho.
A vantagem das provas rotatórias é que o desvio da cúpula pode se produzido por métodos precisos, entretanto o equipamento é caro e sofisticado. A desvantagem é que estimula os dois sistemas simultaneamente não dando informação unilateral. Quanto aos testes calóricos variam no volume de água usada e na temperatura. Um mesmo estímulo é usado para comparar os dois lados evocando todas as conseqüências da inclinação cupular. Neste caso faltando um elemento, uma das sequelas; esta ausência pode ser de grande importância. Por exemplo: havendo nistagmo, vertigem sem náusea, após a estimulação adequada, pode haver indicação de lesão da ponte na parte baixa ou lesão medular com interrupção dos impulsos ao núcleo dorsal eferente do X.
Havendo nistagmo e náusea sem vertigem, pode haver indicação de lesão no tronco cerebral com interrupção de impulsos a centros mais altos onde a vertigem é interpretada.
Havendo vertigem e náuseas sem nistagmo pode indicar uma lesão no tronco cerebral mediano com interrupção de impulsos aos núcleos dos músculos oculares.
O registro eletronistagmográfico permite detectar o nistagmo latente presente com os olhos fechados, velocidade angular da componente lenta e características nistagmicas de lesões cerebrais e periféricas.
Bibiografia
1. ADES, M.W. and ENGSTRÖM, M - 1965- Cit. Lindeman, 1969.
2. ADRIAN, E. D. - Discharges from vestibular receptors in the cat. J. Physiol. (Lond.) 1943, 101: 389-407.
3. ALONSO, C. G. Anatomia y fisiologia del laberinto posterior. Editorial paz Montalvo (Madrid). 1966: 156.
4. BREUER. - 1891- Cit. Huizinga, E. 1955
5. BUYS. - 1924: Cit. Alonso, C. G. (1966)
6. CARLSTRÖM, D., ENGSTRÖM, H. and HJORTH, S.- Electrom microscopic and X-ray differation studies of sttoconia. Laryngoscope. 1953, 63:1052
7. CLARK, B. - The vestibular system. Annual Review of Psychology. 1970, 21: 273-305.
8. CUM BROWN, A. (1874) - Ciit. Huizinga, E. 1955 - March. E. (1875). Cit. Huizinga, E. 1955.
9. DIJKGRAAF, S. - The functioning and significance of the lateralline organs. Biol. Rev. 1963, 38 (1): 51.
10. DOHLMAN, G. F. - 1925 - Cit. Alonso C.G. - 1966.
11. DOHLMAN, G. F., and Anggärd, - 1958 - Cit. Neff, W. D. - 1965.
12. ENGSTRÖM, H. - On the double innervation of the sensory epithelia of the inner ear. Acta Oto-Laryngol. 1958, 49 : 109.
13. ENGSTRÖM, H., Ades, H. W., and Hawkins, J. E. Jr. - Structure and function of the sensory hairs of the inner ear . J. Acout. Soc. Am. -1962, 34 : 1956.
14. EWALD, J. R. - Physiologishe Untersuchungen Über das endorgan nervus octavus, Wiesbaden, Bergman, 1892.
15. FLOCK, K. A. - Electron microscopie and electrophysiological studies on the lateral line canal organ. Acta Oto. Laryng. (Stock.) 1965, Suppl. 199 : 1-90.
16. FLOCK, A. and Wersäll, J. - A study of the orientation of the sensory hairs of the receptor cells in the lateral line organs of fish, with special referente to the function of lhe receptors. J. Cell. Biol. 1962, 15 (1) 19.
17. NEFF, W. D. - Contributions to sensory physiology, Academic Press. New York and London. 1965. 1.
18. OLIVEIRA, J. A. - A Técnica de Preparação de superfície do Epitélio vestibular. 1973. Em preparação.
19. OLIVEIRA, J. A. - Efeitos do Salicilato de Sódio no labirinto da Cobaia. Tese. Faculdade de Medicina de Ribeirão Preto, USP, 1971.
20. PLOTZ, E., and Perlman, H. B. - A Histochemical study de the cochlea. Laryngoscope, 1955, 65 : 291.
21. RASMUSSEN, G. L. - The olivary peduncle and other fiber projections of the superior olivary complex. J. Comp. neurol. 1946, 84: 141.
22. STEINHAUSEN, W. - Über die Beobachtung de cupula in den bogengangsam pullen des labyrinths des lebenden hechts. Arch. Ges. physiol. 1933, 232: 500.
23. TRINCKER, D. - Permanent potenciais in the semicircular canal system of the guinea pig and their changes in experimental cupula leads. Arch. Ges. Physiol. 1957, 264: 351.
24. WERSALL, J. - Studies on the structure and innervation of the sensory epithelium of the cristae ampullaris in the guinea pig. Acta Oto. laryng. 1956. Suppl. 126, 1.
25. VILSTRUP, T. - Studies on the structure and function of the semicircular canais. 1950. Munks gaard. Copenhagem.
26. WERSALL, J. - Crista ampullares. In: submicroscopic structure of the inner ear. (ed. S. lurato) Oxford and London: Pergamon Press. 1967:
195-210.
27. WERSALL, J. - Vestibular receptor celis in fish and mammals. Acta otolayng. 1961. Suppl. 163: 25.
28. HILDING, D., and Wersall, J. - Cho-Laryng. 1962. 55 : 205.
lines and its relation to the nerve endings in the inner ear. Acta Oto
29. HUIZINGA, E. - Anatomie et Physiologie comparées de l'appareil vestibulaire périphérique. Annales d'otolaryngologie, 1955, 4: 250-265.
30. IURATO, S. - Efferent fibtes to the sensory cells of Corti's organ. Expti. Cell. Res. 1962. 27: 162.
31 . KIMURA, R. and Wersall, J. - Termination of the olivo-cochlear bundle in relation to the outer hair celis of the organ of corti in guinea pig. Acta oto-laryng. 1962, 55 : 11.
32. LINDEMAN, H. H. - Studies on the morphology of the sensory regions of the vestibular apparatus. Springer-Verlag Berlin Heidelberg New York, 1969: 1-113.
33. LOWENSTEIN, O. (1966) - Cit. Lindeman, 1969. (1966).
34. LOWENSTEIN, O., Osborne, H. P., and Wersall, J. - Structure and innervation of the sensory epithelia in the labyrinth of the thornback ray (gaja Clavata). Proc .Roy. Soc. 1964, 160: 1.
35. LOWENSTEIN, O. and Roberts, T. D. M. - The equilibrium function of the otolith organs of the thornback ray (gaja Clavata). J. Physiol. (London)m 1950, 110: 392.
36. LOWENSTEIN, O., and Sand, A. The mechanism of the semicircular canal. A study of the responses of single-fibre preparations to angular a acelerations and to rotation at constant speed. Proc. Roy. Soc. (London) - 1940, 129 : 256.
37. LOWENSTEIN, O., and Wersäll, J. A funcional interpretation of the electron microscopic structure of the sensory hairs in the cristae of the elasmobranch Raja Clavata in terms of directional sensitivity. Nature, 1959, 184: 1807.
38. McCABE, B. F. - The quick component of nystagmus: a presentation of a theory of its origin and mechanism involving the dynamic rhytmic inhibition of the slow component based upon a comprehensive Review of previos work and additional experimental evidence. Laryngoscope. .. 1965, 75 : 1619-1646.
Endereço do Autor: Gen. Osório, 362 - 9.º andar Ribeirão Preto - SP.
* Trabalho realizado com auxílio da FAPESP.
** Professor Assistente Doutor da F.F.C.L. - Fisiologia Humana. Professor Assistente Doutor contratado F.M.R.P. (USP) Cadeira de Otorrinolaringologia.


Imprimir: ![]()
All rights reserved - 1933 /
2026
© - Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico Facial