

Ano: 1974 Vol. 40 Ed. 1 - Janeiro - Abril - (2º)
Seção: Artigos Originais
Páginas: 4 a 20
SISTEMA COCLEAR - ESTIMULAÇÃO DAS CÉLULAS CILIADAS (*)
Autor(es): José António A. Oliveira (**)
![]()
As vibrações transmitidas pela cadeia ossicular do ouvido médio à janela oval, pela platina do estribo chegam a perilinfa que circunda e banha o labirinto membranoso que contém os órgãos terminais do equilíbrio e audição.
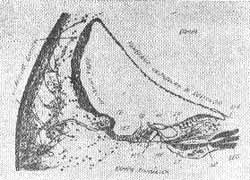
A cóclea tem a forma de uma concha tipo caracol. A parte interna tem uma divisão formando duas escalas paralelas: a inferior é a escala ou rampa timpânica e a superior escala vestibular, as duas contém perilinfa.
A membrana divisória da cóclea é formada por três partes: lâmina espiral que é uma ponte óssea do lado interno enquanto do lado de fora se estende um ligamento grosso conhecido como ligamento espiral. Estas duas estruturas são ligadas pela membrana basilar relativamente fina sobre a qual está o órgão de Corti. Um compartimento em forma de cunha entre as duas escalas é conhecido como duto coclear. É separado da escala vestibular pela membrana de Reissner e da escala timpânica pela membrana basilar (Fig. 1).
O duto coclear contém a endolinfa que parece fluído intracelular. Sua função é nutrir o órgão de Corti. O duto coclear é contínuo, via duto de Hansen com os canais semicirculares.
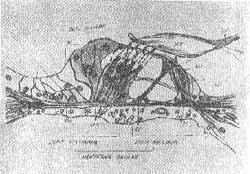
As células ciliadas são arranjadas segmentarmente em fileiras bem definidas. Existe uma fileira de células ciliadas internas e várias fileiras de células ciliadas externas (3 fileiras na espira inferior, 4 fileiras na espira média e 5 fileiras na espira apical).
FIG. 1 - Limites do Duto Coclear - (OLIVEIRA, 1973).
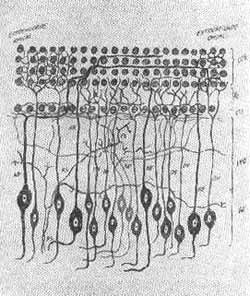
FIG. 2 - Estrutura do órgão de Corti - (OLIVEIRA, 1973).
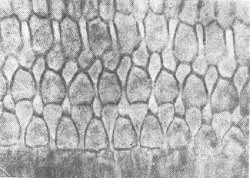
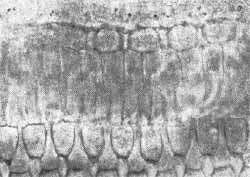
O número total de células internas é cerca de 7.000 e células ciliadas externas 24.000. As células apresentam aproximadamente 40 estereocílios na base e 100 no ápice (Fig. 2, 3, 4, 5). Sobre as células ciliadas está uma estrutura gelatinosa conhecida como membrana tectorial na qual as terminações ciliares estão embebidas. A estrutura da cóclea e órgão de Corti normal e patológico vem sendo estudadas pelas técnicas modernas de preparação de superfície. Em contraste de fase (Fig. 6, 7, 8) Oliveira, Marseillan, 1971) e microscopia eletrônica (Bredberg et als, 1972).
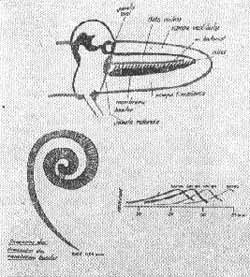
Para se entender a fisiologia do ouvido interno quando estimulado por um som podemos imaginar a cóclea como um tubo reto dividido pela membrana basilar longitudinalmente (Fig. 9). Nota-se que esta membrana se alarga gradualmente a partir do estribo em direção superior, a massa e complacência duas características físicas da membrana também aumentam na mesma direção (Fig. 9).
FIG. 3 - Fotomicrografia com contraste de fases do órgão de Corti normal. Células ciciadas externas (1.400 x).
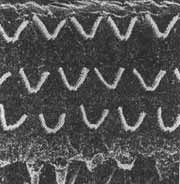
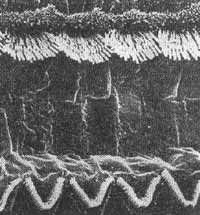
FIG. 4 - Disposição dos cílios nas células ciliadas externas do órgão de Corti - (BREDBERG, 1972).
FIG. 5 - Fotomicrografia em contraste de fases das células ciliadas internas e pilares do órgão de Corti (x 1.400).
FIG. 6 - Disposição dos cílios na fileira de células estiadas internas e primeira fileira de células ciliadas externas - (BREDBERG, 1972).
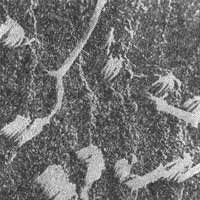
FIG. 7 - Fotomicrografia com contraste de fases do órgão de Corti. Observam-se lesões provocadas por neomicina - (x 1.400).
FIG. 8 - Alterações dos cílios das células sensitivas do órgão de Corti (BREDBERG, 1972).
FIG. 9 - Esquemas indicativos da Fisiologia do ouvido interno.
O estribo vibrando na janela oval, atua como um pistão provocando na rampa vestibular uma onda de pressão que atua em toda a cóclea ao mesmo tempo, devido a velocidade do som e às dimensões pequenas da cóclea.
Esta onda de pressão vai exercer seus efeitos na membrana basilar. Esta membrana na espira basal pode ser leve e rígida acompanha as vibrações da perilinfa, deformando-se sincronicamente com as vibrações do estribo.
Ao longo da membrana basilar, onde a massa vai aumentando as deformações não são sincrônicas com as vibrações do estribo e sim com certo retardo, retardo este que aumenta progressivamente com a distância do ponto considerado ao estribo. Resultante destes fatos forma-se uma onda progressiva (von Békèsy, 1960) (Fig. 9).
Esta onda se propaga na membrana basilar do estribo ao helicotrema. O ponto de máxima amplitude da onda progressiva varia com a freqüência do som estímulo. A área da membrana basilar próxima ao estribo vibra com sons de todas freqüências. A extensão da membrana basilar estimulada é maior quanto mais baixa a freqüência. A membrana se deforma igualmente para freqüências muito baixas.
Como o órgão de Corti está fixo à membrana basilar ele acompanha a mesma em seu padrão de deformação. Com isto os cílios das células receptoras que estão presos a membrana tectoria são deformados por um deslisamento da membrana tectoria em relação à lâmina reticular. Assim as células ciliadas são estimuladas.
As células ciliadas do órgão de Corti não tem kinocílio, mas tem um corpo basal (centríolo) mais próximo a "stria vascularis" do que os estereocílios (Engström et al 1962; Flock et al (1962). De acordo com Békèsy (1953) as células ciliadas são estimuladas por um deslocamento tangencial da membrana tectorial em uma direção radial. Deste modo também o órgão de Corti apresentaria polarização morfológica e funcional (Engström et al 1962; Flock et al 1962; Flock, and Wersall 1963).
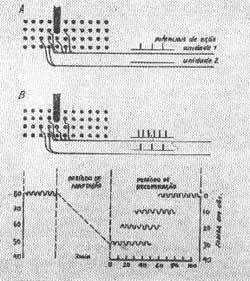
A estimulação das células ciliadas é acompanhada de fenômenos elétricos que podem ser registrados através de eletródios colocados em regiões próximas ao órgão de Corti. Uma técnica simples é colocar um eletródio em contacto com a membrana da janela redonda e um eletródio indiferente. Este método registra a atividade elétrica da espira basal (Simmons, 1962). Com esta técnica observa-se que existem dois tipos de potenciais quando as células são estimuladas: o potencial microfônico coclear que se origina no órgão de Corti (células ciliadas) e que tem as características de um potencial gradativo dos receptores sensoriais, potencial gerador, que reproduz a onda do som que constituiu o estímulo. A função deste potencial é despolarizar as terminações nervosas da via aferente correspondente, iniciando o potencial de ação da fibra que pode ser registrado.
Temos utilizado esta técnica para estudo do efeito de antibióticos do grupo oligossacarídeos (ototóxicos), (Oliveira, Marseillan, 1969), salicilato (Oliveira, 1971), quínino (Cicilini, Oliveira, 1973) (Fig. 10).
Uma aplicação desta técnica no homem é o electrococleograma (Marseillan, Oliveira, 1972). Método audiométrico de importância para avaliação da audição em crianças quando a audiometria tonal liminar não pode ser aplicada.
Consiste num sistema de registro de atividade elétrica coclear no homem.
Usa-se um sistema de geração de clíques para estimular o ouvido em um circuito acústico aberto. A atividade elétrica é recolhida por um eletródio inserido na pele do canal auditivo externo. Os potenciais são ampliaficados, filtrados e analisados por um computador digital de programas fixos. Com esta técnica obtém-se respostas elétricas que consistem de potencial de ação do nervo acústico com duas ondas negativas de amplitude decrescente sendo o potencial microfônico coclear insignificante.
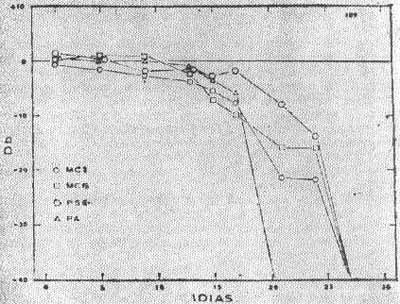
FIG. 10 - Gráfico mostrando, a queda dos Potenciais Elétricos do ouvido interno após administração de 200mg/Kg/dia Paramomicina.
O oitavo nervo craneano, o nervo acústico contém o primeiro neurônio da via auditiva. Este é bipolar e seu corpo está no gânglio espiral no modíolo da cóclea. As terminações destes neurônios fazem sinapses centrais com o núcleo coclear e seus dentritos estão complexamente distribuídos sobre o órgão de Corti.
Suprimento nervoso do órgão de Corti
(Fig. 11)
A penetração dos processos periféricos ou dentríticos das células nervosas do gânglio espiral na papila basilar é acompanhada de uma germinação ativa durante o desenvolvimento embriológico. Deste modo se estabelece uma íntima relação entre as terminações das fibras do nervo coclear e o órgão de Corti. Esta relação está altamente elaborada nas cócleas desenvolvidas do homem e outros vertebrados de boa audição. Esta relação é de suprema importância para a função específica da cóclea e para o receptor periférico da audição.
Os processos dentríticos referidos são dispostos em feixes que se dispõem radialmente ao eixo do modíolo, passando através de canais existentes na lâmina espiral óssea do lado vestibular. Alcançam em seguida a papila espiral basilar ou órgão de Corti do duto coclear membranoso através do lábio vestibular do limbus laminar. Quanto ao arranjo dos terminais nervosos no órgão de Corti, infelizmente, o seu conhecimento é ainda incompleto. A descrição que daremos é baseada em fatos já estabelecidos aparentemente no presente.
FIG. 11 - Inervação do órgão de Corti - (OLIVEIRA, 1973).
Das terminações nervosas da cóclea humana e outros mamíferos existem algumas diretamente relacionadas com o órgão de Corti sendo por isso considerados como fibras nervosas auditivas verdadeiras. Outras são relacionadas indiretamente com o órgão de Corti.
a) Otoneurônios: são os mais numerosos. Alcançam a papila basilar em feixes dispostos radialmente sem grandes desvios. São também chamados fibras radiais ou diretas. Quando penetram na papila basilar seguem um curso reto dando poucos ramos colaterais finos que entram em contato com a superfície inferior das células ciliadas internas Em seguida cruzam o túnel interno e espaço de Nuel e se dividem em vários ramos secundários e terciários produzindo uma arborização terminal restrita. Estas ramificações desta arborização apresentam botões terminais com os quais aderem à superfície inferior e aos lados das cédulas ciliadas externas. Assim cada fibra periférica deste tipo de célula bipolar do gânglio espiral está relacionada a um pequeno grupo de células ciliadas externas e internas.
b) espironeurônios ou fibras espirais: estas fibras são mais espessas que as anteriores. São, entretanto, menos numerosas. A arborização terminal desta fibra fica a uma certa distância do ponto de penetração na papila basilar. O curso é radial, mais ou menos reto do gânglio espiral à papila basilar, em seguida muda abruptamente seu curso de direção para retornar longitudinalmente. Na papila basilar as terminações são encontradas em feixes por entre e abaixo das fileiras de células ciliadas externas. Do mesmo modo que as fibras já descritas, a uma certa distância do ponto de entrada da fibra, na papila basilar, eles terminam em arborização com ramos secundários e terciários. Estes ramos também apresentam botões terminais que tocam a superfície das células ciliadas externas por baixo e ao longo dos lados. Cada fibra espiral liga-se a um grupo de células ciliadas de três, quatro ou mais fileiras, sendo relacionada a poucas células internas através de alguns ramos colaterais vistos no ponto de entrada na papila.
c) platineurônios: fibras espirais internas, fibras difusas: são finas com arborização pequena espalhando sobre um grande segmento da cóclea, e, aparentemente, só terminam nas células ciliadas internas.
As células ciliadas externas são inervadas por ambos os tipos de fibras sendo que cada fibra faz sinapse com muitas células.
Nas células ciliadas internas a inervação é das fibras radiais que fazem sinapses com poucas células receptoras.
O fato de uma fibra nervosa se ligar com múltiplas células ciliadas externas serviria para aumentar a sensibilidade do sistema através da soma spacial. Entretanto a inervação múltipla das células ciliadas internas axiliaria o aumento da zona dinâmica do receptor considerando que as váias fibras aferentes apresentam limiares distintos.
No órgão de Corti bem como em todos os níveis da via auditiva existem terminações eferentes de um sistema que leva informação das estruturas superiores para as inferiores. É importante porque permite que as estruturas centrais exerçam sua regulação sobre os receptores periféricos. Existem várias destas vias cujo conhecimento ainda é escasso. Uma delas foi bem estudada: o feixe-olivo-coclear de Rasmussen do complexo olivar ao órgão de Corti (direto e cruzado). A estimulação (Neff, 1960, Nomoto et al, 1964) deste feixe produz redução na amplitude do potencial de ação do nervo acústico e aumento dos potenciais microfônicos cocleares. A destruição no gato parece afetar a discriminação e a freqüência. Ainda não se conhece o funcionamento exato destas vias.
Fenômenos fisiológicos do sistema coclear
Discriminação de freqüência
Os potenciais de ação percorrendo as fibras nervosas pelas vias auditivas até o cérebro determinariam a discriminação de altura do som pelo sistema nervoso central por dois mecanismos.
1) a distinção dos sons de alta freqüência, se deveria a que cada freqüência estimularia grupos de fibras diferentes isto porque o órgão de Corti seria excitado em uma área relativamente restrita correspondente aos pontos de maior amplitude da onda progressiva.
2) os sons de freqüência baixa põem em vibração grandes extensões da membrana basilar e conseqüentemente ativam um grande número de fibras, a informação quanto a altura seria transmitida pela sincronização das descargas com o som estimulador.
Discriminação de intensidade
Mecanismo de recrutamento de unidades sensitivas.
Aumento da intensidade do estímulo há ativação das células receptoras antes estimuladas por estímulos mais fracos ativando também as células com limiares mais altos que são assim recrutadas. Alguns dos receptores ativados são parte da mesma unidade sensitiva e a freqüência dos impulsos nervosos na unidade, assim, aumenta. Por causa da superposição parcial e interdigitação de uma unidade com outra, também são estimulados receptores de outras unidades e conseqüentemente mais unidades descarregam-se. Deste modo, mais vias aferentes são ativadas e isto é interpretado no cérebro como um aumento em intensidade da sensação (Fig. 12).
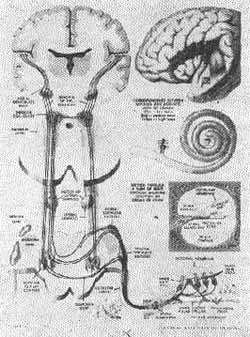
Vias auditivas
(Fig. 13)
No núcleo coclear o agrupamento de unidades de freqüência próxima em áreas circunscritas permite a chamada localização tonotópica.
No complexo olivar superior não foi comprovada localização tonotópica. A este nível o funcionamento é mais complexo.
É a esta altura da via auditiva que se inicia o processo de comparação das informações que vêm dos dois ouvidos permitindo a localização auditiva. Estas informações que vêm do ouvido contralateral chegam através do corpo trapezóide.
FIG. 12 - Esquemas indicando mecanismo de recrutamento e adaptação.
FIG. 13 - Vias auditivas - (MYERS Et AI, 1970).
Colículo inferior - Nestes núcleos as respostas unitárias induzidas por tons puros, alcançam o máximo de complexidade.
Na cavidade neuronal da via auditiva, da cóclea ao colículo inferior, a informação sensorial sofre um processamento crescente, representado pela maior complexidade das respostas, pela maior participação das informações provenientes do ouvido contralateral.
O estudo das respostas na estação seguinte: corpo geniculado medial, por estimulação auditiva são formados por uns poucos potenciais no início da estimulação.
Cortex cerebral auditivo - Escassas respostas à estimulação provocada pelos tons puros.
O que se observa nas lesões corticais é o prejuízo da localização auditiva. (Neff, 1968).
Como conclusão observa-se que existem diversos níveis de análise e integração da informação na via auditiva.
Ao nível mesencefálico se completa provavelmente os mecanismos de discriminação tonal simples.
Ao nível mais superior especialmente o cortex cerebral, teríamos as estruturas que constituiriam os sistemas de decisão e de integração de estímulos complexos da audição direcional e do reconhecimento da linguagem.
A eletroencefaloaudiometria também chamada audiometria cortical, se baseia na alteração que produz um som no traçado normal do EEG. Baseando-se nos conhecimentos anatômicos e fisiológicos do setor neural do aparelho auditivo, sabemos que as vias cocleares a partir do entrecruzamento, levam fibras representativas de ambas as cócleas, para as duas áreas auditivas.
Um eletródio colocado em ou sobre uma região apropriada do cérebro registra atividade elétrica evocada por estimulação acústica ou por estimulação elétrica do nervo auditivo ou de uma pequena região da via auditiva.
Os barbituratos têm sido usados na anestesia pois as transmissões sinápticas nos núcleos auditivos que ligam a cóclea ao cortex cerebral são resistentes aos efeitos depressores destas drogas.
Este método tem provado ser particularmente útil para mapear a extensão e processo de representação central da cóclea. Por contraste o uso de cloralose resulta em um cérebro altamente excitável e experimentos sob estas condições têm revelado foco de potenciais auditivos evocados não vistos sob a ação dos barbituratos.
Potencial evocado
Em animais sob anestesia barbitúrica os fenômenos elétricos que ocorrem no cortex após estimulação de um órgão do sentido podem ser seguidos com um eletródio explorador ligado a outro situado em um ponto indiferente, à certa distância.
Se o eletródio explorador estiver sobre a área receptora primária de um sentido particular aparece uma onda positiva de superfície com uma latência de 5 a
12 ms, seguida de uma pequena onda negativa e, logo de uma deflexão positiva maior e mais prolongada com latência 20 a 80 ms.
A primeira seqüência positiva negativa é o potencial evocado primário e a segunda é a resposta secundária difusa.
A localização da resposta primária é altamente específica e somente: pode ser observada onde terminam as vias provenientes de um órgão particular do sentido. Com efeito ela é tão distinta que foi utilizada para a leitura de mapas das áreas sensitivas do córtex.
A onda positiva provavelmente, representa atividade nas fibras que sobem das camadas corticais profundas para as superfícies e a pequena deflexão negativa, provavelmente representa atividade que se propaga lateralmente na fibra de associação destas camadas superficiais.
Resposta difusa secundária positiva, de superfície é algumas vezes seguida por uma onda negativa ou de uma série de ondas.
A resposta secundária não é localizada aparecendo ao mesmo tempo sobre a maior parte do córtex e outras partes do encéfalo. Não é devida a propagação lateral da resposta evocada primária. Deve ser portanto devida a atividade ascendente da parte abaixo do córtex.
Há evidências de que os impulsos aferentes responsáveis pela resposta difusa secundária sobem pelo tronco cerebral provavelmente pela substância reticular ativadora.
Audiometria eletroencefálica
Usando um computador de soma ou memória pode se estudar as respostas evocadas. Dando-se centenas de estímulos o computador faz a média acusticamente das respostas. São ligados eletródios na cabeça para colher a voltagem do potencial entre o vértex e o lobo da orelha.
O exame é feito com o doente adormecido ou sedado.
Adormecido a tendência é diminuir a resposta. Sedativos como: hidrato de cloral, cloropromazina neubutal e valium não produzem distorsões na forma da onda.
Interpretações dos sinais originados da patologia coclear revelados pelos testes audiométricos, a luz da fisiologia
Em primeiro lugar quando ocorre lesão do sistema coclear ao nível do receptor ou das vias o indivíduo apresentará hipoacusia que será percebida pelo paciente e que poderá ser avaliada por um audiograma tonal liminar ou de tons puros. Por este método de análise podemos determinar se a audição é normal para tons puros de diferentes freqüências para a condução óssea e aérea e determinar o deficit. As enfermidades que provocam este tipo de hipoacusia que chamamos neuro-sensorial geralmente prejudicam mais a audição das freqüências altas.
O audiograma revelará queda simultânea da curva representativa da audição aérea e óssea.
É importante, feito o diagnóstico de uma hipoacusia neuro-sensorial identificar a lesão como sendo das células receptoras ou das fibras nervosas ou seja hipoacusia sensorial ou neural.
Recrutamento
Quando a lesão é coclear o paciente apresenta o recrutamento, estes pacientes não toleram sons intensos.
É um dos fenômenos que permite a discriminação de intensidade de um estímulo, por exemplo, o som.
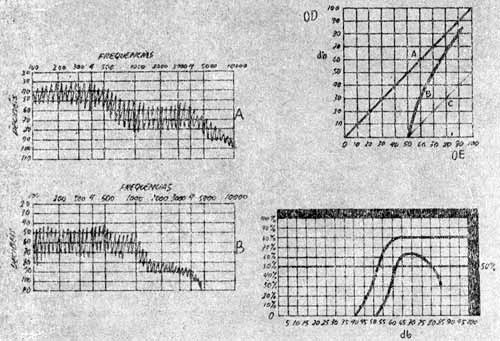
No caso do paciente com surdez devido lesões de receptores no órgão de Corti, quando o estímulo é pequeno a surdez se manifesta, quando se aumenta progressivamente a intensidade de um som se chega normalmente a um certo nível de intensidade supraliminar na qual o ouvido surdo ouve (Fig. 14).
FIG. 14 - Testes audiométricos. A esquerda presença do recrutamento. A figura superior a direita mostrando a curva B, presença de recrutamento figura inferior direita mostra as curvas Logoaudiométricas normal e Patológica.
Deste modo com este estímulo supraliminar, a audição é inteiramente recuperada. Este fenômeno foi descrito por Fowler com o nome de recrutamento. A explicação é a seguinte: com as lesões de células sensitivas que não transmitem impulsos às unidades nervosas a audição diminue. Porém se a intensidade do som aumenta as células receptoras que subsistem e que tem limiares maiores passam a ser estimuladas e enviarão impulsos a um número cada vez maior de unidades nervosas do nervo acústico para o transporte dos estímulos até os centros. Assim o receptor recruta fibras deixadas sem emprego pela destruição do epitélio sensorial do órgão de Corti. A existência deste fenômeno permite diferenciar duas classes de surdez:
a) as devido lesões do ouvido interno e localizadas no epitélio de Corti (Recrutamento +).
b) as que provém de uma afecção radicular do VIII para (recrutamento -).
O recrutamento do ponto de vista audiométrico é uma distorsão de intensidade, devido uma contração do campo audiotonal.
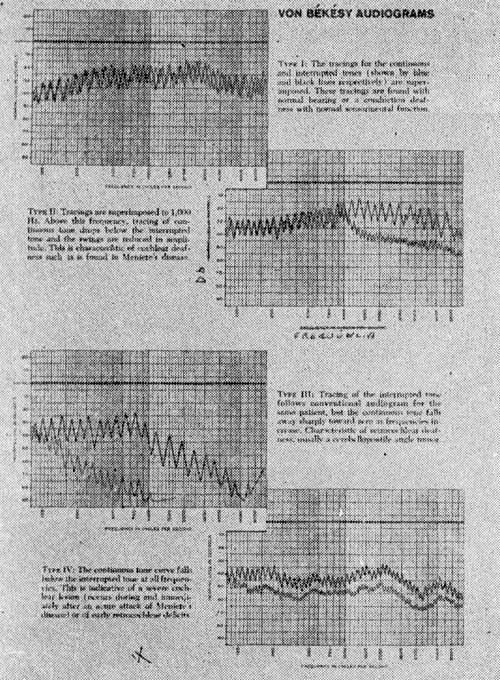
Este fenômeno ou fenômenos conseqüentes ou relacionados podem ser determinados por várias provas audiométricas supra-liminares como Fowler S.I.S., I. Luscher, Rejer, audiometria automática de Békèsy (Fig. 15).
FIG. 15 - Audiometria automática de Von BÉKÉSY. Gráficos III e IV mostrando adaptação alterada. Patologia Retro-Coclear.
Outro fenômeno que está alterado na patologia do sistema coclear é a discriminação da palavra.
As pessoas que tendem a ter perda nas altas freqüências com ou sem recrutamento, têm dificuldade em compreender a palavra por duas razões: as consoantes são de freqüência mais alta e são pronunciadas menos intensamente do que as vogais. Por isso tal paciente escuta a voz mas tem dificuldade em compreender o que está sendo dito. Na realidade a compreensão da fala é mais importante do que qualquer outro aspecto de audição, por isso é importante fazer testes para discriminação da fala. Isto é feito pela audiometria vocal, logoaudiometria (Fig. 14).
Deste modo uma habilidade de discriminação da fala é determinada. Esta habilidade geralmente concorda bem com a curva audiométrica para tons puros se a lesão for no órgão de Corti. Será muito maior a inabilidade se a lesão for retrococlear.
Audiometria direcional é feita para detectar perda de localização sonora que ocorre por lesões nas partes altas das vias auditivas.
Adaptação
(Fig. 12)
A adaptação perestimulatória consiste numa redução gradual na sensação de intensidade produzida pelo estímulo adaptante. Este processo algumas vezes culmina com o desaparecimento completo da percepção. Acompanha o fenômeno a redução da amplitude dos potenciais microfônicos cocleares e da amplitude da resposta neural.
Durante a adaptação haverá também modificação na lateralização da imagem sonora produzida pelo estímulo adaptante sustentado num ouvido e o mesmo estímulo apresentado brevemente ao outro ouvido. Ocorrerá mudança nos impulsos neurais em várias estações ao longo das vias auditivas e em alteração na sensação de altura do estímulo sonoro adaptante e no processo espacial de descarga dos elementos neurais.
Este fenômeno de adaptação estará aumentado nas lesões neurais do sistema coclear, este fato pode ser analisado pelo "tone decay test" e pela audiometria automática de Von Békèsy (Fig. 15).
Audiometria da fala
Dois tipos de medida clínica pode ser obtida. Em outro exemplo o procedimento clínico usual é repetir o paciente os sons que escuta.
Um é a determinação do limiar de recepção da palavra (limiar logoaudiométrico) limiar da intelegibilidade em que o paciente identifica a palavra normal, o outro é o índice de discriminação.
Baseia-se numa avaliação da compreensão da palavra para estabelecer o grau de invalidez social que provoca uma surdez. Para que a compreensão da palavra esteja normal duas funções devem ser normais e discriminação de freqüência do som e a discriminação da intensidade, fenômenos fisiológicos já citados.
Então quando há uma queda aérea e óssea na audiometria tonal laminar estaremos diante de uma patologia do sistema coclear neuro-sensorial, temos que saber do ponto de vista clínico se a hipoacusia é devida a lesão no órgão de Corti, nervo acústico ou central.
Analisando os vários fenômenos fisiológicos já comentados por meio de testes audiométricos especiais podemos ter uma idéia pelo menos aproximada do local lesionado, é claro que analisando-se os dados em conjunto e não separadamente.
Pesquisando-se o recrutamento e a discriminação da palavra podemos tirar conclusões:
Lesões no órgão de Corti
- queda nos agudos abrupta
- presença de recrutamento (distorção subjetiva de sensação de intensidade)
- a compreensão da linguagem diminue após certo limite, devido a distorsão vocal conseqüente ao recrutamento.
- doença de Menière - cocleotoxicoses.
Nervo acústico - lesões radiculares
- ausência de recrutamento
- intelegibilidade se afeta igualmente
- neurinoma acústico, tumor pontocerebeloso, neurite tóxica ou infecciosa, trauma nervoso, lesões degenerativas.
Surdez central
- ausência de recrutamento
- lesão bilateral
- intelegibilidade defeituosa
- transtornos escassos em relação aos sinais neurológicos
- transtornos de localização auditiva.
Bibliografia
BREDBERG, G., ADES, H. W., ENGSTRÖM, H. - Scanning Electron microscopy of the normal and pathologically altered organ of Corti. Inner ear studies. Acta oto-laryngologica. 1972. Suppl. 301: 3.48.
CICILINI, G. A.; OLIVEIRA, J. A. - Efeitos do Quinino oral na coclea da cobaia. Ciência e cultura. 1973. V. 25, 6 : 310.
ENGSTRÖM, H. ADES, H. W. and HAWKINS, J. E., Jr. - Structure and functions of the sensory hairs of the inner ear. J. Acoust. Soc. Am. 1962, 34: 1956-1963.
FLOCK, A., and WERSALL, J. - A study of the orientation of the sensory hairs of the receptor cells in the lateral line organ of fish, with especial reference to the function of the receptors. J. cell. Biol. 1962, 15: 19-27.
FLOCK, A.; KIMURA, R.; LUNDQUIST, P. G. and WERSALL, J. - Morphological basis of directional sensitivity of the outer cells in the organ of Corti. J. Acoust. Soc. Am. 1962, 34 : 1351.
MARSEILLAN, R. F.; OLIVEIRA, J. A. - Registro de atividade elétrica coclear no homem. Aspectos técnicos e resultados preliminares. Revista Brasileira de Otorrinolaringologia. 1972, 38 (2) : 185 :188.
MYERS, D.; SCHLOSSER, W.; WOLFSON, J. R.; WINCHESTER, R. A.; CARMEL, N. H. - Otologic diagnosis and the treatment of deafness. CIBA. 1970. Vol. 22, N. 2: 1-68.
NEFF, W. D. - Role of the auditory cortex in sound discrimination. In: Neural Mechanisms of the Auditory and Vestibular Systems, Charles Thomas, Springfield, 1960: 211.
NEFF, W. D. - Localization and lateralization of sound in space. In: Hearing Mechanisms in Vertebrates, Churchill, London, 1968: 207.
NOMOTO, M.; SUGA, N.; KATSUKI, Y. - Discharge pattem and inhibition of primary auditory nerve fibers in the monkey J. Neurophysiol., 1964, 27: 768. OLIVEIRA, J. A. - Efeito do salicilato de sódio sobre o labirinto da cobaia, Tese, Faculdade de Medicina de Ribeirão Preto, USP, 1971, Ribeirão Preto. OLIVEIRA, J. A. - Citoarquitetura coclear. Revista Brasileira de Otorrinolaringologia. 1973. Vol. 39(1) : 12-23.
OLIVEIRA, J. A.; MARSEILLAN, R. F. - Efeitos da neomicina e da paromicina sobre o labirinto anterior da cobaia. Anais XVIII Congres. Bras. Otorrinolaringol., Tomo I, Porto Alegre, 1969 : 201.
OLIVEIRA, J. A. MARSEILLAN, R. F. - Estudo de lesões cocleares pela técnica de preparações de superfície na cobaia. Revista Brasileira de otorrinolaringologia. 1971. Vol. 37(1): 1-5.
SIMMONS, F. B.; BEATTY, D. L. - The significante of round window-recorded cochlear potentials in hearing. An autocorrelated study in the cat. - Ann. Otol. (St. Louis), 1962, 71 : 767.
VON BÉKÉSY, G. - Shearing microphonics produced by vibrations wear the inner ear and outer hair cells. J. Acoust, Soc. Amer. 1953, 25: 786-790.
VON BÉKÉSY, G. - Experimental models of the cochlea with and without nerve supply. Vestibular Systems, Charles Thomas, Springfields, 1960:3.
(*) Trabalho realizado com auxílio da FAPESP
(**) Professor Assistente Doutor da F.F.C.L. - Fisiologia Humana. Professor Assistente Doutor contratado F.M.R.P. (USP) Cadeira de Otorrinolaringologia.

Imprimir: ![]()
All rights reserved - 1933 /
2026
© - Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico Facial