

Ano: 1993 Vol. 59 Ed. 4 - Outubro - Dezembro - (1º)
Seção: Artigos Originais
Páginas: 236 a 248
O MECANISMO ELETROBIOMECÂNICO ATIVO DA CÓCLEA.
Cochlear bioelectrical active process.
Autor(es): José Antonio A. de Oliveira*
Palavras-chave: transdução coclear, discriminação coclear, amplificação coclear, emissões acústicas, fisiologia coclear, células ciliadas, eletromotilidade
Keywords: cochlear transduciton, cochlear discrimination, cochlear amplification, otoacustic emissions, cochlear physiology, hair cells, electromotility
Resumo:
Inicialmente é descrita a cóclea e suas estruturas internas. O mecanismo de transmissão passivo através das ondas progressivas é revisto. Uma descrição estrutural e fisiológica do sistema de células ciliadas externas e internas é realizada com ênfase nas diferenças entre os dois sistemas. Enfoque é dado à capacidade de contração das células ciliadas externas e a de codificação da mensagem sonora das células ciliadas internas.
Os papéis da inervação aferente e eferente das células ciliadas externas e internas são ressaltados, especialmente suas ações no processo eletrobiomecânico das células ciliadas externas. O papel deste processo é aumentar a capacidade de amplificação coclear e tornar mais perfeita a seletividade freqüência da membrana basilar.
É enfatizado pelos canais fônicos na estimulação das células ciliadas externas e internas. Finalmente são descritas as 3 etapas da transdução coclear, base da fisiologia da cóclea.
Abstract:
This paper describes the inner cochlear structure. The passive mechanism of progressive waves is revised A structural and physiological description is done with emphasis in differences of outer and inner hair cells systems, and the capacity of contraction of the outer hair cells as well the codification of the sound message of inner hair cells.
The importance of afferent and efferent innervation of outer and inner hair cells are stressed specially their actions in the electrical biomechanical process of the outer hair cells. The importance of this process is to increase the capacity of cochlear amplification. It is emphatized the outer and inner hair cells stimulation. Finally, the three steps of the cochlear transduciton, are describe.
![]()
CÓCLEA (figura 1)
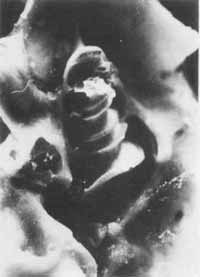
A cóclea ou caracol constitui o labirinto anterior que faz parte do ouvido interna ou labirinto. Suas paredes são ósseas, limitando três tubos enrolados em espiral em torno de um osso chamado columela ou modíolo, ao redor do qual dão duas voltas e meia. Para melhor entendermos a anatomia e fisiologia coclear, podemos imaginar a cóclea desenrolada. Sua base que é mais alargada apresenta as duas janelas, oval e redonda. Os três tubos coleares têm disposição paralela, e são de cima para baixo: a rampa (escala) vestibular que se limita com o ouvido médio pela janela oval; a rampa (escala) média ou canal coclear que contém o órgão de Corti; a rampa (escala) timpânica que se limita com o ouvido médio pela janela redonda. As duas rampas, vestibular e timpânica, comuniquem-se pelo helicotrema no ápice da cóclea e contêm perilinfa que apresenta composição química semelhante á do líquido extracelular (maior concentração de íons sódio). O canal coclear apresenta endolinfa com composição semelhante à do líquido intracelular (maior concentração de íons potássio). Como podemos observar a rampa vestibular está separada do canal coclear pela membrana vestibular de Reissnner e o canal coclear está separado da rampa timpânica pela membrana basilar, onde está o órgão de Corti. Esta membrana alarga-se da sua base (0,04 mm) até o ápice (0,4 mm), aumentando, portanto, sua massa. Sobre o órgão de Corti está a membrana tectorial com sua borda interna fixa ao modíolo e sua borda externa livre.
TRANSMISSÃO DA ONDA SONORA NO OUVIDO - ONDAS PROGRESSIVAS
O som que é transmitido pelo sistema timpano-ossicular provoca vibrações do estribo que penetram na perilinfa da escala vestibular, em nível da janela oval. O movimento do estribo para dentro e para fora determina a transmissão da vibração pela perilinfa e esta se move em direção à rampa timpânica e canal coclear, variando a pressão na janela redonda (janela de descompressão). Durante a transmissão das vibrações pela perilinfa, ocorrem deslocamentos simultâneos das membranas do canal coclear, da membrana de Reissnner e da membrana basilar. Deste modo, inicia-se uma onde de oscilação na base da membrana basilar que vai propagar-se, em direção ao helicotrema, até uma certa extensão da mesma, dependendo da freqüência sonora. Estas oscilações da membrana basilar, para cima e para baixo, têm amplitude em torno de 10-11 mm. Como a membrana basilar é mais estreita na base junto ao estribo e vai se alargando em direção ao ápice da cóclea, suas características físicas como elasticidade, rigidez, massa se alteram ao longo da membrana. Daí a maior vibração da membrana diferentemente ao longo do seu comprimento.
FIGURA 1 - Cóclea da cobaia aberta, mostrando as espiras cocleares com órgão de Corti.
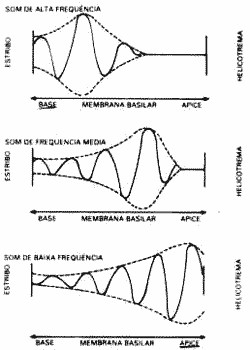
FIGURA 2 - Propagação da onda viajante na membrana basilar.
Esta onda propagada "viajante' tem um ponto de deflexão máxima na membrana basilar entre a origem da onda, junto ao estribo, até onde ela termina. Este local de deflexão máxima corresponde à porção da membrana basilar que tem freqüência ressonante natural para a freqüência sonora correspondente. Neste local a membrana vibra com facilidade e a energia da onda se dissipa. A onda termina neste local. Para cada freqüência sonora, este máximo ocorre em áreas diferentes da membrana basilar. Quanto maior a freqüência, mais próximo do estribo. Quanto menor a freqüência, mais próximo do helicotrema. Assim, um som agudo provoca uma onda que viaja por uma distância muito pequena na membrana basilar antes de alcançar a sua oscilação máxima e desaparecer. Com som de freqüência média a onda viaja metade da distância antes de desaparecer. Com som grave a onda viaja ao longo de toda a membrana basilar até alcançar a deflexão máxima próxima ao helicotrema. A existência de locais de oscilações máximas, localiza cada freqüência sonora numa área determinada da membrana basilar. Nestas áreas as células sensoriais são mais excitadas. Assim, cada freqüência provoca excitação máxima em células sensoriais diferentes de áreas diferentes que informam regiões corticais diferentes, podendo ocorrer a discriminação da freqüência sonora (Oliveira, 1971; Schmidt, 1978). Este mecanismo de discriminação é passivo não sendo por isso muito acurado (Fig.2).
ESTRUTURA DOS RECEPTORES - ÓRGÃO DE CORTI
É a estrutura receptora auditiva, formada da basicamente por células de sustentação e células receptoras secundárias ciliadas (estereocílios). Estas células estão situadas sobre a membrana basilar em toda a sua extensão. No órgão de Corti existe uma membrana chamada tectorial, presa à lâmina espiral óssea do modíolo. Esta membrana cobre os cílios das células ciliadas externas entrando em íntimo contato com eles durante as vibrações da membrana basilar. Nos pólos inferiores das células ciliadas encontramos sinapses com neurônios que apresentam seus corpos celulares nos gânglios espirais de Corti, localizados na cóclea. Os axônios destes neurônios constituem o nervo coclear. O fornecimento de energia para a cóclea e a manutenção da composição dos líquidos cocleares é realizado pela stria vascularis, região onde se concentram vasos sangüíneos (Oliveira, 1973) (Figs.3,4).
Uma técnica que é muito utilizada para estudo da estrutura histológica do órgão de Corti é a técnica de preparações de superfície, na qual o órgão de Corti como um todo é analisado com microscopia de contraste de fases ou microscopia eletrônica de varredura (Oliveira, 1971; 1974; 1989)
FIGURA 3 - Órgão de Corti de um gato. Microscopia óptica (De Engstrom, B. & Engstrom, H. Audiology, 11:6, 1973. In. - OLIVEIRA, J.A.A. Audiovestibular Toxicity of Drugs. CRC Press. Boca Raton. U.S.A., 1989).
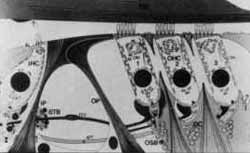
FIGURA 4 - Estrutura do órgão de Corti TM - membrana tectorial; IHC - célula ciliada interna; IP - pilar interno; OP - pilar externo; DC - Deiter's cens; STB -feixe espiral do túnel; EFF -fibras eferentes; RFF -fibras aferentes; OSB -feixe espiral externo - OHC - células ciliadas externas (De Engstrom H. and Engstrom B. The Ear, Widex. Dinamarca, 1988).
CÓCLEA ATIVA
Os conceitos sabre a fisiologia eoclear nos últimos 10 anos se modificaram fundamentalmente. Vários setores de pesquisa permitiram novos avanços como amicrofisiologia, neurohistoquímica neuroquímica e cultura de células, contribuindo significativamente para os conhecimentos do funcionamento coclear.
No órgão de Corti existem dois sistemas de células ciliadas. O sistema de células ciliadas externas e o sistema de células ciliadas internas (Brownell, 1985; 1990). Até recentemente os estudiosos de anatomia e fisiologia coclear tinham observado diferenças anatômicas e fisiológicas entre as células ciliadas dos dois sistemas sem saber exatamente as implicações das mesmas na fisiologia coclear (Figs. de 3 a 8).
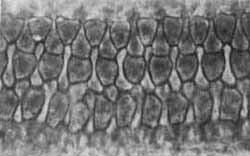
FIGURA 5 - Células ciliadas do órgão de Corti de cóclea normal. Observam-se três fileiras de células externas e uma fileira de células ciliadas internas. Fotomicrografia em contraste de fases.
Estrutura do sistema de células ciliadas externas
O sistema de células ciliadas externas apresenta 10000 a 14000 células no homem. Estas células têm a forma cilíndrica, e estão dispostas em três fileiras ao longo das espiras cocleares. São menores na base da cóclea e maiores no ápice. Estão sobre a membrana basilar do canal coclear, possuindo uma firme ligação com esta membrana. As células ciliadas externas são banhadas pela endolinfa no pólo ciliar e pela perilinfa nas partes laterais. A endolinfa só banha o pólo ciliar. Estas células são envolvidas parcialmente pelas células de sustentação, ficando os espaços de Nuel entre elas permitindo movimento das mesmas. No pólo basal há ligação firme das células ciliadas externas com as células suporte de Deiters que estão ancoradas sobre a membrana basilar, permitindo que os movimentos das células ciliadas externas possam repercutir sobre a membrana basilar e estrutura do canal coclear.
Em relação aos estereocílios, cada célula apresenta dezenas de cílios em três fileiras, mantendo uma disposição padrão semelhante à letra W. Os esterocifios mantêm-se ligados entre si em cada fileira através de ligamentos formando uma unidade. Os cílios das três fileiras são de alturas diferentes, sendo os mais internos mais curtos, os mais externos mais longos e implantados na membrana tectorial.
FIGURA 6 - Células ciliadas externas do órgão de Corti de cóclea de cobaia normal. Fotomicrografia em contraste de fases.
FIGURA 7 - Microscopia eletrônica de varredura de órgão de Corti de cóclea de cobaia normal (De M. Lenoir. In: Pujol. R. Cochlear Physiology and Pathophysiology: Recent data Drugs roday, 26, 43, 1990.)
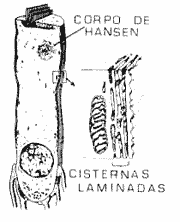
Cada cílio mais interno de cada fileira, mais curto, está ligado ao cílio mais longo da camada seguinte através de ligamento de elastina que sai do pólo apical do cílio mais curto. Os estereocfilos mais longos são acoplados à membrana tectorial que fica superiormente localizada no órgão de Corti. O padrão W é mais aberto nas células ciliadas da espiral basal. Recentemente foi demonstrada a presença de proteínas contráteis nas células ciliadas externas como acima, miosina, tropomiosina e agentes reguladores como a calmodulina (Flock e cols., 1986). Outra estrutura que tem sido muito estudada atualmente é o sistema de cisternas laminadas que só existem nas células externas e é formado por vários planos de membranas logo abaixo da membrana lateral da célula (Brownell, 1990) (Fig. 9).
O sistema de cisternas laminadas é um conjunto de sacos ou vesículas membranosas, achatados, adjacentes à membrana plasmática lateral. O complexo praticamente só existe nas células ciliadas externas ao longo de todo o comprimento destas células entre a placa cuticular do pólo superior e o núcleo no pólo inferior, na mesma área das células expostas aos espaços de Nuel. Este sistema pode ajudar a manter a forma da célula, devido à ausência de uma rede de citoesqueleto desenvolvido, nas células ciliadas externas. A ausência de citoesqueleto bem definido nestas células é compensada por uma pressão hidrostática positiva, mantida provavelmente pelo alto conteúdo de glicogênio destas células, que mantém o citoplasma hiperosmótico. O complexo de cisternas laminadas citado pode manter a necessária integridade estrutural destas células, sendo a força elástica deste complexo, evidente (Brownell, 1990).
Fisiologia do sistema de células ciliadas externas
Do ponto de vista fisiológico o potencial elétrico de repouso das células ciliadas externas é de -70mV. Descobertas recentes, nesta última década têm acrescentado grandes novidades no conhecimento do funcionamento das células ciliadas externas, que vieram revolucionar o entendimento da fisiologia coclear e da audição.
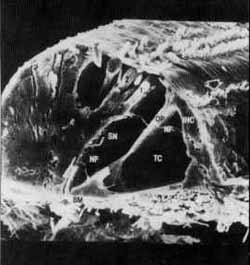
FIGURA 8 - O órgão de Corti humano em microscopia eletrônica de varredura (S.E.M.): 1.2.3 -fileiras de células ciliadas externas, IHC -fileira de células ciliadas internas, setas - cílios das células ciliadas externas e das células ciliadas internas, OP -pilar externo do túnel de Corti; NF -fibras nervosas; TC - túnel de Corti; SN - espaço de Nuel; BM - membrana basilar. (De Engstrom, H. anã Hengstrom, B. The Ear Widex, Dinamarca 1988).
Foi evidenciado que estas células não têm capacidade de atuar como receptor coclear não codificando a mensagem sonora. Têm capacidade de dois tipos de contração, rápida e lenta, sendo efetores cocleares ativos devido à eletromotilidade, ou seja, suas propriedades biomecânicas (Brownell, 1985; Zenner e cols., 1985).
Seriam estas células, devido à energia mecânica liberada na contração rápida, responsáveis pelas otoemissões acústicas. Estas são respostas de energia de audiofreqüência da cóclea com origem nas células ciliadas externas, que podem ser captadas por um microfone miniatura sensível, quando se aplicam estímulos acústicos como cliques, no canal auditivo externo. Esta energia liberada na cóclea é transmitida pela cadeia ossicular e membrana do tímpano ao meato auditivo externo, onde pode ser registrada. Estas respostas são devidas a um biomecanismo ativo das células ciliadas externas. As contrações rápidas induzidas eletricamente, ocorrem na ausência de ATP, presença de baixos níveis de cálcio, refutando um papel das proteínas contráteis (Brownell,1985; Zenner, 1985).
Às otoemissões são registráveis com limiar normal ou perda auditiva inferior a 30dB. A intensidade das otoemissões cresce com a intensidade da estimulação até 20 ou 30 dB, onde existe certa linearidade da resposta. Após este limite ocorre saturação da resposta deixando de ser linear. O limiar de detecção das otoemissões chega a ser mais baixo que o próprio limiar subjetivo, em aproximadamente 10 dB. Podem ser registradas também no meato auditivo externo, otoemissões acústicas espontâneas, ou seja, sem estimulação, em 40% de indivíduos com audição normal. Estas otoemissões são controladas pelo sistema eferente mediano cruzado que chega às células ciliadas externas, alterando as contrações rápidas através de contrações lentas, tipo muscular (activa e miosina), o mediador liberado sendo a acetilcolina. O registro das otoemissões acústicas vem sendo utilizado para triagem em crianças de risco de perda auditiva, sendo um método objetivo, fácil, rápido.
FIGURA 9 - Sistema de cisterna laminadas abaixo da membrana das células çiliadas externas (De Brownell, W E. - Outer hair cell eletromotility and otoacoustic emissions, Ear and Hearing, 11(2). 12, 1990.
Como vimos, do ponto de vista fisiológico, as células ciliadas externas teriam capacidade de contração rápida e lenta, ou seja, propriedades eletrobiomecânicas. Estudos têm demonstrado que a contração rápida não requer ATP e cálcio e segue ciclo por ciclo a freqüência de estimulação até várias dezenas de KHz. Deste modo, o sistema de células ciliadas externas funcionaria como um amplificador coclear e seria capaz de acurada seletividade frequencial.
A contração rápida seria a base do mecanismo ativo induzido pelo deslocamento dos estereocílios, sendo como vimos o suporte das otoemissões acústicas. O mecanismo da contração rápida ainda não está definido, mas parece estar relacionado à função das cisternas laminares que ficam sob a membrana plasmática celular. Seria um processo de eletrosmose (Brownell, 1990).
Os elementos do citoesqueleto das células ciliadas externas estão localizados na placa cuticular e estereocílios onde existe a activa e outras proteínas similares e na região sinóptica ou infranuclear. Como já vimos, uma característica destas células é a falta de elementos esqueléticos na região axial, entre o núcleo e a placa cuticular onde não existem cadeias longas de moléculas protéicas. As respostas eletromóveis rápidas manifestam-se nesta região da célula, onde estão as cisternas laminadas. A eletrosmose poderia ser um mecanismo para explicar a transdução eletromecânica, com capacidade de gerar força mecânica rápida, em resposta aos potenciais microfônicos cocleares.
A interface líquido-sólida, entre o citoplasma e as membranas do sistema de cisternas laminadas provê um substrato adequado para eletrosmose. A eletrosmose é o movimento de fluído induzido por um campo elétrico adjacente a uma superfície carregada eletricamente (membrana celular) e resulta de uma migração eletroforética de íons próxima à superfície de sua carga. A velocidade do fluído é proporcional ao campo elétrico aplicado. O movimento do fluído e o campo elétrico na eletrosmose são paralelos à superfície e não através dela, sendo este movimento paralelo ao maior eixo da célula, determinando alterações no seu comprimento e diâmetro (contração rápida) (Brownell, 1990).
As células ciliadas externas tornam a cóclea um verdadeiro amplificador mecânico que permite o aumento de até 50 dB da intensidade de um estimulo, pois provocaaumento na amplitude da vibração da membranabasilar, permitindo aumento da estimulação das células ciliadas internas, cujos estereocilios normalmente não estariam em contato com a membrana tectorial. Como a contração rápida segue ciclo por ciclo a freqüência de estimulação até dezenas de KHz, ocorre sensível modulação na discriminação de freqüência da cóclea, tendo grande implicação na seletividade freqüência desta estrutura, o que não poderia ser explicado somente pelo mecanismo passivo das ondas progressivas de Von Békésy (Békésy, 1960). Resumindo, as propriedades eletrobiomecânica das células ciliadas externas ou o mecanismo ativo coclear tem grande implicação na estimulação amplificada das células ciliadas internas para a codificação da mensagem sonora e a capacidade de discriminação de freqüências.
A contratação lenta das células ciliadas externas seria similar à contração muscular pois foi demonstrada a presença de proteínas contráteis como ativa e miosina nestas células, bem como reserva de cálcio (Zenner e cols., 1985). A contração lenta das células ciliadas externas induzida por ATP é inibida pela citocalasina B, bifosfato inorgânico, trifluorperazina, isto indicando um mecanismo móvel dependente da ativa modulado pela calmodulina que pode ativamente ajustar as propriedades mecânicas das células ciliadas externas e da membrana basilar durante a estimulação auditiva. Este tipo de contração lenta, tipo muscular, é possível devido à presença de ativa, miosina, alfa-actinina, fimbrina e tropomiosina nas células ciliadas externas. As contrações lentas não seguem ciclo por ciclo a freqüência de estimulação sonora, isto podendo ocorrer até 1 KHz, portanto não possibilitam a seletividade de freqüências acima de 1KHz. Estas contrações são moduladas pelo sistema eferente medial. Estas contrações controlariam a tonicidade das células ciliadas externas e regulariam as propriedades mecânicas da membrana basilar.
O sistema eferente medial das células ciliadas externas provocaria o mecanismo das contrações lentas que modularia as contrações rápidas, tendo este sistema ativo implicações audiológicas importantes como: a capacidade do indivíduo de detectar um sinal no ruído; afinamento da seletividade freqüência; proteção contra superestimulação acústica, focalização de atenção para um fenômeno acústico; regulação da amplificação coclear, funcionando como um amortecedor, durante a amplificação para melhor captação do estímulo sonoro pelas células ciliadas internas. O sistema eferente medial, quando estimulado pode também reduzir as otoemissões acústicas, regulando as contrações lentas com atenuação das contrações rápidas.
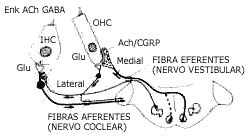
FIGURA 10 - Esquema das sistemas aferentes e eferentes das células ciliadas externas e internas com os prováveis neurotransmissores nas sinopses; ACh = acetilcolina, GABA - ácido gama-aminobutírico, ENK - encefalina, GLU - glutamato, CGRP - peptídeo relacionado ao gen calcitonina. (De Pujol. Drugs of Today, 26: 43, 1990).
Inervação do sistema de células ciliadas externas (Fig.10)
Inervação aferente
Os neurônios são pouco numerosos, ramificados, constituindo 5% das fibras do nervo auditivo. São neurônios do Tipo 11, com fibras espirais não mielinizadas e ramificadas (Gold, 1948). Enviam aos centros nervosos exclusivamente mensagens lentas (fibras finas), pouco seletivas (fibras ramificadas e provavelmente não auditivas). Atualmente nenhuma resposta à estimulação sonora pode ser registrada nestas fibras aferentes.
Existe uma fibra nervosa para cada 10 ou 20 células ciliadas externas. Não se conhece ainda o neurotransmissor da sinapse entre a célula ciliada externa e o neurônio aferente. Estas sinapses não respondem à estimulação sonora. Como hipótese, este sistema captaria informações sobre o estado de contração das células ciliadas externas, não conduzindo nem codificando mensagens sonoras para o sistema nervoso central. As informações citadas seriam importantes para regular o tono das células ciliadas externas e regular as contrações rápidas destas células, através das contrações lentas. Outra hipótese seria funcionar como sistema de alerta na presença de ruído intenso.
Inervação eferente
As células ciliadas externas possuem o sistema eferente medial e lateral. O sistema eferente medial apresenta fibras mielinizadas com trajeto radial, cada fibra mantendo contato com 15 a 30 células. Sua origem é na região medial do complexo olivar superior no núcleo ventromedial do corpo trapezóide no tronco cerebral. Este sistema é contralateral (70%) e ipsilateral (30%). As sinapses são axossomáticas e o principal neurotransmissor é a acetilcolina e em menor escala o CGRP (peptídeo relacionado ao gen calcitonina).
A estimulação deste sistema eferente medial eleva o limiar das respostas do nervo acústico; reduz as espostas intracelulares das células ciliadas internas; reforça o mascaramento ipsilateral; diminui o traumatismo acústico contralateral. Uma hipótese de funcionamento seria a regulação da contração lenta das células ciliadas externas, atenuando as contrações rápidas, ou seja, o amplificador coclear, funcionando como um amortecedor do amplificados (Helmholtz, 1863). Isto através da regulação do comprimento, tensão, rigidez da células ciladas externas.
Sistema de células ciliadas internas
Existem aproximadamente 3500 células ciliadas internas, são piriformes com estereocílios com a forma do padrão V muito aberto, dispondo-se em uma só fileira sobre a membrana basilar. São bem presas às células suportes bem envolvidas por estas, não ficando espaços livres ao seu redor e banhadas só pela endolinfa no pólo ciliar. Os cílios destas células não alcançam a membrana tectorial e também estão ligados entre si como foi descrito para as células ciliadas externas.
Fisiologia das células ciliadas internas
O potencial de repouso destas células é de 40mV. São transdutores sensoriais. São os verdadeiros receptores da mensagem sonora produzindo codificação em mensagem elétrica, que seria enviada pelas vias nervosas aos centros auditivos do lobo temporal. As curvas de freqüência destas células são semelhantes as do nervo auditivo, ou sejam têm seletividade em freqüência fina (Sellick e cols., 1982). Estas constatações foram feitas através de registros intracelulares das fibras aferentes (Russel e cols., 1977).
Estas células apresentam canais catiônicos, especialmente em nível dos estereocilios na porção apical destes, que são abertos por ocasião da vibração da membrana basilar, quando os cílios são inclinados ao contatar a membrana tectorial. Deste modo haveria entrada de potássio nos canais catiônicos, que provocaria a despolarização das células com conseqüente formação de potenciais elétricos receptores.
Inervação do sistema de células ciliadas internas
Inervação aferente
São neurônios ganglionares do Tipo I, constituindo 90 a 95% das fibras do nervo acústico. São fibras radiais sem ramificações. Existem 10 neurônios Tipo I para cada célula ciliada interna, sendo o neurotransmissor sinóptico o glutamato. Este sistema aferente levaria amensagem sonora codificada para os centros auditivos. O glutamato é tóxico para as células ciliadas internas quando acumulado, ocorrendo lesões nestas células, este poderia constituir um dos mecanismo da presbiacusia (Pujol e cols., 1986).
Sistema eferente
É chamado de sistema eferente lateral e se origina no bulbo ao nível da oliva superior, sendo ipsi lateral com fibras não mielinizadas. Os neuro transmissores mais importantes são: acetilcolina, GABA (ácido -gama- aminobutírico), dopamina, neuropeptídeos (encefalinas, dinorfinas) e CGRP. Não se sabe a exata função deste sistema, talvez inibição durante exposição a ruído intenso (Pujol e cols., 1986).
FISIOLOGIA DA CÓCLEA ATIVA
Após os experimentos de Békésy, durante muito tempo admitiu-se que a membrana basilar vibrava passivamente com a vibração mecânica dos líquidos cocleares, ocorrendo então o mecanismo das ondas viajantes, proposto pelo pesquisador, que seria por sua vez, responsável pela discriminação de freqüência da cóclea ao nível do receptor auditivo periférico (as células ciliadas do órgão de Corti). Entretanto, alguns pesquisadores descreveram em 1958 e 1965 a curva auditiva que mostrava a freqüência específica das fibras do nervo acústico o que permitia a grande seletividade freqüência destas fibras e suas características tonotópicas (Katsuki e cols., 1958, Kiang e cols.,1965). A curva de reposta de uma fibra do nervo acústico em função da intensidade e da freqüência do som, mostra uma freqüência própria à cada fibra. Ao redor desta freqüência e a sensibilidade e a seletividade freqüencial da fibra são máximas. A melhor freqüência (freqüência característica) depende do seu ponto de origem na membrana basilar. A curva neural é muito mais fina que a curva de resposta mecânica da membrana.
Estas evidências trouxeram uma grande dúvida. Como pode ocorrer discrepância entre a tonotopia mais grosseira, uma seletividade de freqüência imprecisa, explicada pelas ondas viajantes (curva mecânica de Békésy) e a tonotopia específica, apurada das fibras do nervo acústico? Esta controvérsia foi esclarecida com algumas descobertas realizadas recentemente. Em 1978, foi demonstrado que transdutor neurosenssorial, que codificava a mensagem sonora era apenas o sistema de células ciliadas internas, sendo as células ciliadas externas incapazes de atuar como receptores da mensagem sonora. Este estudo foi feito com registros intracelulares das fibras aferentes das células ciliadas internas após estimulação acústica (Russel e col., 1977). Em 1988 foi demonstrado que as células ciliadas internas têm seletividade em freqüência fina como a das fibras do nervo auditivo (Sellick e cols., 1978). Em 1982 foi demonstrado que as propriedades mecânicas da cóclea são muito mais finas do que as que Békésy descreveu (Sellick e cols., 1982). Além disso, outros autores afirmaram que a seletividade da membrana basilar é incomparável a das fibras do nervo acústico (Russel e cols., 1977; Sellick e cols., 1982).
Com estas descobertas concluímos que a seletividade de freqüência da membrana basilar e das células ciliadas internas são correspondentes, ou seja, muito específicas e acuradas, semelhantes ao das fibras nervosas auditivas. Por outro lado, desde 1969, se sabia que havia pequena inervação aferente das células ciliadas externas e extensa inervação aferente das células ciliadas internas, o que poderia explicar, em parte, a importância das células ciliadas internas como os únicos receptores e codificadores da mensagem sonora na cóclea (Spoendlin, 1969).
A discrepância inicial havia sido elucidada, mas qual seria o mecanismo que permitiria à cóclea ou à membrana basilar a sua acurada seletividade de freqüências sonoras?
Já em 1863, Hehmholtz propunha a existência de ressoadores mecânicos específicos para cada freqüência sonora. Gold, em 1948, sugeriu um sistema de amplificação mecânica na cóclea e previu a existência das otoemissões produzidas por este amplificador mecânico.
Este sistema de amplificação mecânica realizaria ao mesmo tempo a amplificação dos sons muitos fracos, sua análise freqüencial fina e o seu amortecimento rápido. Evans, em 1975, sugeriu a existência de um mecanismo eletrobiomecânico que seria um segundo filtro analisador de freqüências além do proposto por Békésy.
Em 1978, Kemp registrou, pela primeira vez, as otoemissões acústicas previstas por Gold e sugeriu que estas emissões seriam produzidas por um mecanismo bioativo coclear. Davis, 1983, afirmou que deveria existir um amplificador de fenômenos mecânicos na cóclea propondo que seriam as células ciliadas externas.
Deste modo, vários autores propuseram a existência de um mecanismo eletrobiomecânico na cóclea, que explicaria a amplificação coclear e a estimulação das células internas bem como a cura da seletividade de freqüência da membrana basilar semelhante e das fibras do nervo acústico.
Finalmente por volta de 1985, vários autores conseguiram provocar contração das células ciliadas externas isoladas em meio de cultura, aplicando no meio estímulos elétricos e químicos (Ashmore, 1987; Brownell, 1984; Zenner e cols., 1985) (Figs. 11, 12, 13, 14).
Deste modo estava demonstrada a presença e funcionamento do amplificador coclear previsto por vários autores. Seria o sistema de células ciliadas externas, com sua capacidade mecânica de contração. Estas células, através deste mecanismo eletrobiomecânico especialmente as contrações rápidas, já explicadas anteriormente, provocariam amplificação e acurada análise freqüencial dos sons. Esta amplificação podendo chegar a 50 dB HL, o que implicaria numa grande amplificação da vibração da membrana basilar, o que provocaria maior sensibilidade das células ciliadas internas que assim poderiam ter seus estereocílios estimulados, alcançando as suas extremidades apicais, a membrana tectorial.
Por este mecanismo, um pequeno número de células ciliadas externas seriam estimuladas pela freqüência específica de um som, permitindo uma acurada discriminação de freqüências.
Resumindo, as células ciliadas externas constituiriam o amplificador coclear, sendo importantes no mecanismo de amplificação do estímulo para determinar o funcionamento das células internas, que seriam as unidades receptoras e codificadoras cocleares e teriam papel importante na seletividade freqüencial da cóclea (Fig.15).
Após a descoberta deste amplificador biomecânico ativo podemos entender as diferenças fisiológicas da cóclea ativa com a cóclea passiva de Békésy e também as diferenças morfológicas já apresentadas entre os sistemas de células ciliadas externas e internas do órgão de Corti, que explicam as grandes diferenças fisiológicas destes dois sistemas.
Etapas da fisiologia da cóclea ativa
PRIMEIRA ETAPA - TRANSDUÇAO MECANOELÉTRICA NAS CÉLULAS CILIADAS EXTERNAS
Potencial de repouso endococlear
É a diferença de potencial existente entre a endolinfa e a perilinfa e de valor de +80mV, com positividade na parte interna do canal coclear e negatividade na rampa vestibular. Acredita-se que este potencial seja gerado pela difusão de potássio da estria vascular para o canal coclear.
Papel do potencial endococlear
A endolinfa banha os extremos superiores das células ciliadas. Como as células ciliadas têm potência de membrana negativo (-40 a -70mV), em relação à perilinfa haverá um potencial de -110 a - 150mV em relação à endolinfa, na superfície superior junto aos cílios. Este elevado potencial elétrico torna a sensibilidade das células ciliadas muito grande, capaz de despolarizar sua membrana com pequenos deslocamentos ciliares.
Estimulação das células ciliadas externas
As reações moleculares que participam na transdução mecanoelétrica das células ciliadas sensoriais estão ainda mal conhecidas. Nos últimos anos apareceram técnicas de isolamento e manutenção das células ciliadas em meio de cultura in vitro. Com esta técnica é possível pesquisar o comportamento, mecânico, elétrico e bioquímico das células ciliadas em diferentes condições de estimulação, podendo, assim, serem esclarecidas as bases do mecanismo de transdução mecanossensorial (Dulon e cols., 1990).
FIGURA 11 - Célula ciliada externa isolada do órgão de Corti de cobaia, em meio de cultura.
FIGURA 12 - Equipamento para estudo e registro da estrutura e contrações das células ciliadas externas em meio de cultura (microscópio invertido, câmara de vídeo, monitor de TV, vídeo)
FIGURA 13 - Célula ciliada externa em repousa (A) e contraída (B).
FIGURA 14 - Célula ciliada externa isolada em estágio de deterioração. Nota-se abaulamento próximo ao núcleo no pólo inferior.
As células ciliadas têm um pólo apical na placa cuticular onde existem os estereocílios e um pólo basal onde estão as sinapses nervosas. O conjunto de cílios está em contato com a endolinfa rica em K + (150mM) e pobre em sódio (1mM). O corpo celular das células ciliadas, especialmente externas, está em contato com a perilinfa que apresenta concentração alta em Na + (170MM) e pobre em K+ (5mM). Como entre a endolinfa e o meio intracelular existe um potencial de -70mV aproximadamente, ocorrerá uma diferença de potencial da ordem de 150mV através da membrana dos cílios e a lâmina cuticular.
Os estereocílios em número de algumas centenas e cilíndricos constituem os receptores das vibrações mecânicas. A deflexão dos cílios em direção ao corpúsculo basal produz uma despolarização excitatória da célula no sentido inverso uma polarização inibitória.
Os potenciais receptores celulares que serão formados podem ser registrados por um eletrodo intracelular in vivo ou in vitro. A resposta elétrica da célula ciliada é função da freqüência e da intensidade da estimulação.
FIGURA 15 - Envelope da vibração da membrana basilar para íon som de 10 KHz. Próximo do limiar o componente relacionado ao mecanismo ativo CCE, desviado em direção ao ápice, provoca um ganho de cerca de 50 dB no limiar e uma filtração muito seletiva. Quando o mecanismo ativo está ausente, somente a curva passiva persiste. (De Davis, 1983. In: Pujol R.: Cochlear Physiology and Pathophysiology: Recent Data Drugs of Today, 26: 43, 1990).
Os estereocílios são extensões da membrana e são rígidos devido uma rede de filamentos de actina, fimbrina, alfa actina, tropomiosina. Tem diâmetro de 500 a 900 mm na parte superior a 100 mm na parte basal. São interligados por filamentos que unem os cílios de cada fileira entre si e com os cílios das outras fileiras. Os primeiros são os ligamentos transversais e os segundos os ligamentos apicais, mais finos que ligam o ápice dos cílios mais curtos a região subapical do cílio mais longo adjacente. Este conjunto relativamente rígido se desloca em conjunto durante a vibração sonora e os filamentos de conexão na extremidade dos cílios poderiam estar relacionados a abertura e fechamento dos canais fônicos de transdução (Dulon e cols., 1990). Embora não exista prova bioquímica acredita-se que os canais de transdução estariam ao nível dos cílios.
O número de canais fônicos ao nível dos cílios seria pequeno havendo sugestão de um único canal por estereocílio e estaria localizado no pólo apical do cílio. Os ligamentos interciliares estariam diretamente relacionados aos canais fônicos de transdução provocando sua abertura e fechamento. Seriam os elementos mecânicos que provocariam a abertura dos canais (Hudspeth, 1989). Outros autores acham que os canais de transdução estariam nas regiões basais dos estereocílios (Ohmori, 1988). Outra teoria relata que o movimento do tufo ciliar produziria um estiramento da membrana dos cílios. Este estiramento poderia ser a origem da abertura de um canal ativado por estiramento (Sachs, 1988). De qualquer modo as células ciliadas externas possuem canais de potássio que seriam abertos pela inclinação dos cílios durante a estimulação acústica, ocorrendo pelos canais catiônicos entrada de potássio e despolarização celular. O fechamento dos canais estaria relacionadas aos íons cálcio que poderiam entrar na célula por canais de cálcio.
Poderíamos descrever o processo de transdução do seguinte modo. O som determinaria a oscilação da membrana basilar alternadamente em direção a rampa superior e inferior da cóclea. A membrana tectorial vai se deslocar com padrão diferente da membrana basilar. Estes deslocamentos da membrana tectorial e basilar determinarão o aparecimento de forças tangenciais sobre os cílios das células ciliadas externas provocando inclinação dos mesmos, pois estes cílios estão em contato com membrana tectorial.
Esta inclinação dos cílios determina a abertura dos canais de potássio com conseqüente entrada de potássio na célula (Fig. 16). Esta despolarização determina o aparecimento de fenômenos elétricos nas células ciliadas, que podemos registrar com eletrodos implantados nas proximidades da cóclea, são os potenciais receptores microfônicos e potencial de somação cocleares.
Como vimos, a vibração sonora da membrana basilar provoca uma voltagem alternante da mesma freqüência, que se superpõe ao potencial de repouso endococlear. Esta voltagem alternante; despolarização, hiperpolarização constitui os potenciais microfônicos cocleares. Estes potenciais comportam-se como microfones, reproduzindo as flutuações de pressão sonora. Suas características são: ausência de período refratário, ausência de latência, ausência de fadiga e limiar elevado. Outro potencial coclear que aparece durante a transdução é o potencial de somação que é um potencial receptor.
SEGUNDA ETAPA - TRANSDUÇÃO ELETROMECÂNICA NAS CÉLULAS CILIADAS EXTERNAS
Após a formação dos potencias nas células ciliadas externas elas se contraem em fase cote a freqüência do som, que provocou o estímulo (contrações rápidas). Como o acoplamento da membrana tectorial com a membrana basilar é mantido pelas próprias células ciliadas externas, ocorre uma amplificação da vibração, numa zona limitada da membrana basilar com máxima efetividade. Neste ponto é que podemos observar o mecanismo ativo coclear devido à capacidade de contração das células ciliadas externas. Deste modo, os potenciais receptores ocorrem quando os cílios são inclinados, resultando da modulação de uma corrente estável pela estria vascular que mantém a concentração de potássio. A modulação mecanoelétrica é mediada, então, pelo estereocílio que provoca o aparecimento dos potenciais elétricos e que elegem mudanças de comprimento eletricamente evocadas das células ciliadas externas em freqüência acústicas determinadas. Assim as células ciliadas externas têm um mecanismo de transdução bidirecional. Convertem energia elétrica acústica (mecânica) em energia neural (elétrica) sendo esta a transdução mecanoelétrica. Estas células possuem também capacidade efetora, pois sinais eletrofisiológicos (potenciais elétricos) aplicados às células são traduzidos em geração de forças mecânicas e alterações no comprimento das células. Esta é a transdução eletromecânica. A conversão de energia em ambas as direções (transdução bidirecional) parece ser única na audição e determina que o ouvido possa produzir sons.
Suporte para relação entre amotilidade da célula ciliada externa e os potenciais receptores COCLEARES seria o fato que, as otoemissões acústicas, que são devido a esta motilidade rápida continuam após a morte, portanto tempo, quanto o potencial endococlear e os microfones cocleares possam ser ainda registrados.
TERCEIRA ETAPA - TRANSDUÇÃO MECANOELÉTRICA NAS CÉLULAS CILIADAS INTERNAS
Estas contrações localizadas das células ciliadas externas, amplificando as vibrações da membrana basilar e tectorial, permitem a estimulação dos cílios mais longos das células ciliadas internas que normalmente não alcançam e não estão acoplados à membrana tectorial (Pujol, 1989). A amplificação de uma área limitada da membrana basilar estimula poucas células ciliadas internas.
FIGURA 16 - Esterocílio em repouso (esquerdo) e em estado de excitação (direito). (De Pujol, R.: Cochlear Physiology anil Pathophysiology: Recent Data. Drugs of Today, 26: 43, 1990).
Às células internas com a inclinação dos cílios são estimuladas. Os canais de K+ são abertos e a entrada destes íons provoca despolarização celular de modo semelhante ao das células ciliadas externas. Quando isto ocorre, na região sináptica há liberação do neurotransmissor e o sistema de enervação aferente leva a mensagem sonora codificada pela célula ciliada interna, na forma de impulsos elétricos (potenciais de ação) às áreas auditivas centrais.
Devemos ressaltar mais uma vez o papel das células ciliadas externas, com suas propriedades contráteis (mecanismo bioativo) possibilitando por amplificação das vibrações intracocleares estimulação das células ciliadas internas. Por outro lado, pela capacidade de estimular poucas células ciliadas internas ou mesmo uma, conforme a freqüência do som, na cóclea, duas freqüências muito próximas excitam de modo diferente duas células internas vizinhas e conseqüentemente estas células enviam mensagens discretamente diferentes ao sistema nervoso central. Esta seria a base para a acurada discriminação de freqüência (Pujol, 1989).
Como bem afirma Aran, as descobertas recente mostram um caminho novo para nossa compreensão sobre os problemas de audição. Assim os distúrbios de discriminação podem estar relacionados a alterações no funcionamento do analisador coclear. Os novos conhecimentos poderão contribuir nas gerações de próteses acústicas futuras ou de próteses da orelha interna, como os implantes cocleares. Também estes progressos no conhecimento da fisiologia coclear poderão no futuro ajudar na prevenção de problemas auditivos de origem periférica.
RESUMO
Resumindo, poderíamos dividir a fisiologia coclear em três etapas fundamentais.
PRIMEIRA ETAPA - TRANSDUÇÃO MECANOELÉTRICA NAS CÉLULAS CILIADAS EXTERNAS
As vibrações mecânicas da membrana basilar e órgão de Corti provocadas pelas vibrações da perilinfa, determinariam deslocamento dos estereocílios das células ciliadas externas acoplados á membrana tectorial. Neste mecanismo de vibração há uma seletividade de freqüências imprecisa caracterizada por mecanismos passivos ligados às propriedades físicas da membrana basilar.
Quanto maior a freqüência do som estimulante o máximo de ressonância se desloca para a base da membrana próxima ao estribo. Os estereocilios deslocando-se cora as vibrações são estimulados abrindo os canais depotássio. Estes íons penetram nas células ciliadas externas provocando o aparecimento de potenciais elétricos receptores corno os potenciais microfânicos cocleares (Figs. 17 e 18).
SEGUNDA ETAPA - TRANSDUÇÃO ELETROMECÂNICA (ATIVAS) NAS CÉLULAS CILIADAS EXTERNAS
Os potenciais elétricos formados provocariam contrações mecânicas rápidas das células ciliadas externas. Estas contrações constituem a base da eletromotilidade e ocorrem em fase com a freqüência sonora estimulante. Estas contrações determinariam uma amplificação da vibração da membrana basilar numa área restrita do órgão de Corti, devido ao acoplamento que as células ciliadas externas realizam entre a membrana basilar e a membrana tectorial. As contrações rápidas poderiam ocorrer por um mecanismo de eletrosmose que depende da presença do sistema de cisternas laminadas nas células ciliadas externas. Este mecanismo constitui a base de funcionamento do amplificador coclear ativo (Fig. 19).
TERCEIRA ETAPA - TRANSDUÇÃO ELETROMECÂNICA NAS CÉLULAS CILIADAS INTERNAS
A amplificação das vibrações da membrana basilar pelo mecanismo ativo das células ciliadas externas provocaria o contato dos cílios mais longos das células ciliadas internas com a membrana tectorial com conseqüente inclinação dos mesmos. Isto nas células de uma área delimitada pequena onde é liberada a energia pelo mecanismo ativo. Nesta área, um pequeno número de células ciliadas internas é estimulado com máxima intensidade. A inclinação e estimulação dos cílios determinaria a despolarização das células ciliadas internas, com formação de potenciais receptores pela entrada de potássio pelos canais iônicos dos cílios. Em seguida há liberação dos neurotansmissores e a formação de uma mensagem sonora codificada em impulsos elétricos, que é transmitida ao sistema nervoso central pelo nervo acústico. A base da acurada discriminação de freqüências é a exitação seletiva de células ciliadas internas muito próximas, que pelas suas enervações próprias enviarão mensagens com pequenas diferenças ao sistema nervoso central (Fig. 20),
FIGURA 17 - Orgão de Corti em repouso. A - Célula ciliada extenua; B - Célula ciliada interna. (De PujoA R.: Cochlear Physiology and Pathophysiology. Recent Data. Drugs of Today, 26: 43, 1990).
FIGURA 18 - Primeira Etapa - Excitação e transdução mecanoelétrica nas células ciliadas externas. (De Pujo/, R.: Cochelear Physiology and Pathophysiology: Recent Data. Drugs of Today, 26: 43, 1990).
FIGURA 19 - Segunda etapa - Contração e transdução eletromecânica das células ciliadas externas. (De Pujo4 R.: Cochlear Physiology and Pathophysiology: Recent Data Drugs of Today, 26: 43, 1990).
FIGURA 20 - Terceira etapa - Excitação e transdução mecanoelétrica nas células ciliadas internas. (De Pujo4 R.: Cochlear Physiology and Pathophysiology: Recent Data Drugs of Today, 26: 43, 1990).
REFERÊNCIAS BIBLIOGRÁFICAS
ARAN, J.M.: L'amplificateur cochléaire. Medicine-sciences, 6:740,1990.
ASHMORE, J.F.: A fast motile response in guinea-pig outer hair cells. The celular basis of the cochlear amplifier. J. Physiol. (London), 388: 323, 1987.
BÉKÉSY, VON H,: Experiments in Hearing. New York, Me Gram Hif, 1960.
BROWNELL, W.E.: Microscopic observation of cochlear hair cell motility. Scann Electr. Microsc, III, 1401, 1984.
BROWNELL, W.E.: Outer hair cell electromotility and otoacoustic emissions, Ear and Hearing, 11(2): 82, 1990.
BROWNELL, W.E.: Outer hair cell electromotility and otoacustic emissions. Ear and Hearing, 11(2):12,1990.
BROWNELL, W.E.; BADER, C.R; BERTRAND,D.; RIBAUPIERRE, Y.: Evoked mechanical responses of isolated cochlear outer hair cells. Science, 227- 194, 1985.
DAVIS, H.: An active process in cochlear mechanics. Hear. Res., 9:79, 1983.
DULON, D. & ARAN, J.M.: Aspects cellulaires et moléculaires de la transduction mécano-elétrique dan f oreille interne. Medicine-Sciences, 6: 744, 1990.
EVANS, E.F.: Normal and abnormal functioning of the cochlear nerve. In: BENCH, RJ., PYE, A.; PYE, S.D.: Sound Reception in Mammals. Londres, Academic Press, 1975 p. 133.
FLOCK, A., FLOCK, B.; ULFENDAHL.: Mechanisms of movement in outer hair cells and a possible structure basis. Arch. Oto rhinol, 243: 83, 1986.
GOLD, T.: The physical basis of the action of the cochlea. Proc. Soc. London B,; 1948 p. 135.
HELMHOLTZ, H.L.F.: Die lehre von den tonempfindungen als physiologische grund lage fun die, Theoric der Musik. 1° ed, vieweg - verlag Brunswick, Allemagne, 1863.
HUDSPETH, A.J.: How the ear's works. Nature, 341: 397, 1989.
KATSUKI, Y.; SUMI, T.; UCHIYAMA, H.; WATANABE, I.: Electric responses of auditory neuronsin catto sound stimulation. J Neurophysiol., 21: 569, 1958.
KEMP, D.T.: Stimulated acoustic emissions from the human auditory System. J Acoust. Soc. Am. 64: 1386, 1978
MANO, N.Y.S.; WATANABE, T.; THOMAS, E.F.; CLARK, L.C.: Discharge patterns of single fibers in the cat's auditory nerve. Research Monograph. Cambridge: Mit Press, 1965 p. 35.
OHMORI, H.: Mechanical stimulation and Fura-2 fluorescence in the hair bundle of dissociated hair cells of the chick. J. Physiol (London), 399: 155, 1988.
OLIVEIRA, J.A.A.: Anatomofisiologia animal comparada. Ribeirão Preto. Editora Universitária, 1971.
OLIVEIRA, J.A.A.: Estudo das lesões cocleares pela técnica de preparações de superfície na cobaia. Rev. Bras. Otorrinol., 37: 1, 1971.
OLIVEIRA, J.A.A.: Citoarquitetura coclear. Rev. Bras. Otorrinol., 39: 12, 1973.
PUJOL, R.: Anatomie et physiologie de la cochleé. Archiv. Internat. Physiol. Bioch., 97 (4): 51, 1989.
PUJOL, R. &LENOIR, M.: The four types of synapses in the organ of Corti. In: ALTSCHCTLER, R.A.; BOBBIN, R.P.; HOFFMAN, D.W. Neurobiology of Hearing. The Cochlea. Raven Press. New York, 1986 p. 161.
RUSSEL, I.J. & SELLICK, P.M.: Tuning properties of cochlear hair cells. Nature, 267: 858, 1977.
SACHS, F.: Mechanical transduction in biological systems. CRC Critical Review in Biomedical Enginneering, 16: 141, 1988. SCHMIDT, R.F.: Fundaments of Sensory Physiology. Berlin, Spring Verlag, 1978.
SELLICK, P.M.; PATUZZI, R.; JOHNSTONE, B.M.: Measurement of basilar membrane motion in guinea-pig using Mossbauer Technique. J. Acoust. Soc. Am. 72: 131, 1982.
SELLICK, P.M. & RUSSEL, LJ.: Intracellular studies of cochlear hair cells: filling the gap between basilar membrane membrane mechanics and neural excitation. In NAUTON, R.F.; FERNANDEZ, A.C. Evoked Electrical Activity in theAuditory Nervous System, New York, Academic Press, 1978 p. 113.
SPOENDLIN, M.: Innervation patterns in the organ of Corti of the cat. Acta Ototaryngol. (Stockh), 67: 239, 1969.
ZENNER, H.P.; ZIMMERMANN, U.; SCHIMITT, V.: Resersible contraction of isolated mammalian cochlear hair cells. Hear. Res., 18: 127, 1985.
* Professor Titular, Chefe do Departamento de Oftalmologia e Otorrinolaringologia da Faculdade de Medicina de Ribeirão Preto - USP.
Endereço: Faculdade de Medicina de Ribeirão Preto - USP. - Departamento de Oftalmologia e Otorrinolaringologia - Av. Bandeirantes, 3900 - 14049-900 - Ribeirão Preto/ SP.












Imprimir: ![]()