

Ano: 2000 Vol. 66 Ed. 1 - Janeiro - Fevereiro - (8º)
Seção: Artigos Originais
Páginas: 52 a 57
A Lei de Wagner-Grossman é Aplicável nas Paralisias da Laringe?
Is The Wagner-Grossman's Law Aplicable to Larynx Paralysis?
Autor(es):
Rui C. M. Mamede*,
Francisco V. M. Filho**,
Bernardo M. Entschev***.
Palavras-chave: paralisia da laringe, laringe, cirurgia experimental da laringe
Keywords: larynx paralysis, larynx, experimental surgery of larynx
Resumo:
Objetivos: O presente trabalho teve como objetivo determinar os fatores que podem influenciar a posição assumida pela prega vocal (PV), em casos de paralisia dos nervos laríngeos recorrente (NLR) e superior (NLS). Material e método: Para tanto, foram utilizados ratos, nos quais se provocou a paralisia da PV de acordo com os seguintes procedimentos: o Grupo A: realizados cortes no NLR em alturas diferentes, em 20 ratos; Grupo B: realizados cortes no NLR e de fibras do músculo cricotireóideo, em 15 ratos; o Grupo C: realizado corte do NLR, obedecendo o movimento da PV no ciclo respiratório, em 7 ratos. Discussão: Observou-se gire a prega vocal permanecia em posição pára-mediana (PPM) na grande maioria dos casos, mesmo quando as fibras do músculo cricotireóideo foram seccionadas ou o músculo foi denervado. Nesta última situação, observou-se que em 20% dos ratos, a PV se mantinha em posição lateral (PO. Nos casos em que o corte do NLR se fazia segundo o ciclo respiratório, a posição da prega era PPM, mesmo quando o corte ocorria em inspiração. Neste caso, a PV, então abduzida, se desloca rapidamente para a linha mediana, como se estivesse sendo puxada. Conclusão: Conclui-se que, nos ratos, a posição da PV independe da reinervação, da sincinesia e da inervação residual, principalmente do músculo cricotireóideo. O movimento da PV quando da secção do NLR durante a inspiração sugere a ação de forças elásticas.
Abstract:
Aim: This study aims to determine the influencing factors in the positioning of the vocal fold (VF) in cases of paralysis of the recurrent and superior laryngeal nerves (RLN, SLN). Material and methods: VF paralysis was caused in rats according to the following procedures: Group A: 20 rats were cut at different sites of the RLN. Group B: 15 rats were cut at the R1.N and cricothyroid muscle fibers. Group C: 7 rats had the RLN cutoff, according to the VF movelnent in the breathing cycle. Discussion: In most cases, the Ws were still in paraimedian position even when the cricothyroid muscle fibers were se sectioned or the muscle denervated. In 20% of the rats, which underwent muscle denervation, the VF's were still laterally positioned. In those cases, when the RLN's were cut following the breathing cycle, the vocal fold was in paramedian position, even when the cut was made in the inspiratoy phase of the breathing cycle. In this case, the VF, then in abdution, quickly moves to the median line, as in was being, pulled. Conclusion: We conclude that the VF position in rats is independent of 1. the cricothyroid muscle action; 2. the site of the lesion in the RLN; and 3. phase of the breathing cycle in which the lesion occurs.
![]()
INTRODUÇÃO
Os sintomas apresentados pelos pacientes que apresentam paralisia da laringe variam consideravelmente. Esta variação de sintomas é observada de paciente para paciente; e até mesmo, com o passar do tempo, num mesmo paciente. Ela é decorrente de posicionamentos diferentes da prega, vocal (PV) em cada tipo de paralisia e também da instabilidade dessa prega, numa mesma paralisia. É conhecido que as paralisias unilaterais geralmente se manifestam com graus variáveis de incompetência glótica, e que em 30% dos casos os pacientes são totalmente assintomáticos.
Através dos anos, observa-se muita discussão entre os autores sobre os fatores que controlariam a posição da PV nas paralisias. O debate ganhou força frente a duas conflitantes hipóteses sobre os responsáveis pelas variações de posição da PV. A hipótese de Semon, também chamada de Semon Rosenbach, descrita primeiramente por Semon e, algum tempo depois, por Rosenbach, afirma que as fibras nervosas da musculatura abdutora da laringe são mais sensíveis a danos que as fibras correspondentes da musculatura adutora; então, urna lesão nervosa lentamente progressiva faria a PV assumir inicialmente uma posição medial e, mais tarde, lateral. Todavia, os casos clínicos raramente apóiam a hipótese de Semon- Rosenbach; e, contrariamente, com o tempo, a posição observada da PV é de movimento lateral para medial.
A hipótese de Wagner-Grossman afirma que, na paralisia do nervo recorrente, a PV se mantem próxima à linha média pela ação cio músculo cricotireóideo, que é inervado pelo NLS e tem função adutora. Segundo esta teoria, a lesão bilateral do nervo laríngeo recorrente (NLR) deveria causar obstrução das vias aéreas, pela presença das pregas vocais próximas ã linha média, obstrução esta que deveria ser aliviada nas paralisias do músculo cricotireóideo. Dedo, em 1970, ao estudar a paralisia da laringe em cachorros e humanos, comprovou a hipótese ele Wagner-Grossman. Alguns métodos de tratamento da paralisia do nervo laringeo recorrente são baseados neste fato, ou seja, que a perda da ação do músculo cricotireóideo resultaria na lateralização da PV afetada. Mas, apesar dos relatos iniciais afirmarem que a inativação do músculo cricotireóideo produzia um alívio da obstrução do fluxo aéreo, as aplicações clínicas subseqüentes falharam na sua confirmação. Existem vários casos relatados que não se alinham na hipótese de Wagner-Grossman; entre eles, citam-se os de Woodson, 1993, a, b5, 6, baseados em eletromiografias.
Diante de tais indefinições, procuramos neste trabalho identificar, em ratos, os fatores que possam influencia na posição assumidas pelas PV nos casos de paralisia vocal.
MATERIAL E MÉTODO
A proposta desse trabalho tem como objetivo determinar fatores que possam influenciar no posicionamento da PV paralisada. Para tanto, foram provocadas em animais situações de paralisia das PV por secção dos nervos vago, laríngeo recorrente e laríngeo superior. Estes cortes foram realizados em diferentes alturas dos nervos vago e recorrente, com o objetivo de determinar pontos que pudessem produzir paralisias com a PV em posições diferentes, pois sabe-se que o nervo laríngeo recorrente tem fibras nervosas que estimulam tanto Os músculos abdutores quanto os adutores da laringe.
Para simular uma situação de paralisia do nervo laríngeo superior, procurou-se ora destruir o músculo cricotireóideo, ora seccionar as fibras do NLS.
Foi realizado um plano piloto com dois cães, dois ratos e um gato. Na proposta inicial, os sujeitos em estudo seriam cães, visto que são os animais cuja anatomia apresenta-se com muita semelhança à do homem, além de ter um tamanho superior a outros animais comumente usados em cirurgia experimental. No teste piloto feito com os cães, observou-se a impossibilidade de identificação do posicionamento da PV, visto que a anestesia geral provoca diminuição acentuada do movimento da mesma. Convém ressaltar que a mobilidade da PV é condição primordial para a identificação da posição que a mesma adquire quando há secção da sua inervação.
Uma outra forma de se definir o posicionamento ela PV é através da observação dos sinais clínicos decorrentes de uma paralisia das cordas vocais. No entanto, esta tentativa foi frustrante, especialmente quando as pregas vocais permaneceram em posição para mediana (MIM), pois provocou asfixia e morte do animal, especialmente nos cães após terem seu NLR seccionado. Diante destas dificuldades, novas tentativas foram feitas com Outros animais, como ratos e gatos. Com o rato, a experiência foi bem sucedida; no entanto, outras desvantagens se apresentaram, devido às dificuldades de manuseio do nervo laríngeo recorrente, por conta de sua pequena dimensão, além do freqüente espasmo da laringe, provavelmente devido ao trauma provocado pelo laringoscópio.
Por conta disso, experimentou-se a utilização do gato como sujeito a ser investigado, especialmente por seu tamanho ser superior ao do rato. Porém, tal animal apresentou os mesmos problemas oferecidos pelos cães quando anestesiados, ou seja, a movimentação das pregas vocais estava diminuída, necessitando, portanto, da observação dos sinais clínicos no pós-operatório, o que lhe causou a morte por asfixia horas após o experimento.
Diante desses fatos, optou-se pela utilização de ratos, pois foi o animal que apresentou maior mobilidade das PV acompanhando os movimentos respiratórios, depois de anestesiados. Este fato permitiu a filmagem desses movimentos e detectar através de vídeo o momento exato da paralisia e a posição da PV. Para tanto, foram usados 33 ratos hamster, de ambos os sexos, pesando cada um 350 gramas, em média.
Os ratos foram submetidos à tricotomia da face anterior do pescoço, da região esternal, até a mandibular, previamente à intervenção.
Utilizamos diferentes técnicas anestésicas, ora com evaporação de clorofórmio, ora com injeção de clorofórmio subcutâneo, ora com quelicin endovenoso.
Uma incisão longitudinal, na linha mediana anterior do pescoço, era aplicada da mandíbula até a fúrcula esternal, com bisturi lâmina 12. Procedia-se a dissecção, procurando afastar os elementos anatômicos da linha mediana para a exposição do conduto laringo-traqueal. Esta dissecção era realizada com tesoura e bisturi. Identificação rigorosa do nervo laríngeo recorrente era realizada bilateralmente. Ao mesmo tempo, introduzia pela boca do animal, uma lente tipo Hopkins, de 0° grau, de 3 mm de espessura e 25 cm de comprimento, que era acoplada à televisão e ao vídeo, através de uma micro-câmera. Todo o exame era filmado e armazenado em fitas.
O estudo foi realizado através de três tipos de experimento, a saber:
Grupo A: procedia-se o corte do nervo NLR, em alturas diferentes em relação à borda inferior da cartilagem cricóide dos ratos.
Grupo B: cortavam-se as fibras do músculo cricotireóideo antes de se proceder ao corte do NLR dos ratos. Em alguns desses animais, ao invés de se cortar o músculo, cortava-se o nervo laríngeo superior (NLS)
Grupo C: o corte do nervo dos ratos era efetuado, seguindo o posicionamento da PV no ciclo respiratório.
Para a realização dos experimentos, utilizou-se instrumental cirúrgico, na grande maioria delicado, constituído por: bisturi, tesoura, pinças hemostáticas, afastadores, fios de sutura, além de luvas, máscaras e toucas.
RESULTADOS
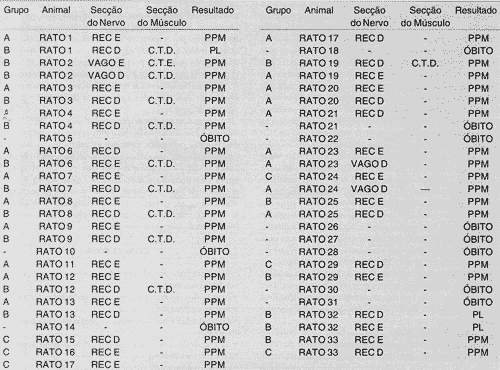
Utilizamos 33 ratos, sendo que 10 faleceram antes que qualquer procedimento pudesse ser finalizado. Em quatro, o óbito ocorreu após termos realizado o experimento em um dos lados. Assim, em 19 o procedimento realizado foi duplo, ou seja, bilateralmente.
Grupo A: Utilizaram-se de 20 ratos para se proceder às técnicas descritas para este grupo. O nervo laríngeo recorrente esquerdo foi seccionados em 13 situações, o direito o foi por cinco vezes e o vago durante duas vezes. Em todas estas situações verificou-se que a PV permanecia parada em posição pára-mediana (PPM). Não houve alteração na posição da PV quando se variava a altura elo corte, em relação á borda inferior da cartilagem cricóide, nem mesmo quando o corte era aplicado no nervo vago.
Grupo I3: a lesão do nervo NLR era acompanhada de secção das fibras do músculo cricotireóideo. Esta técnica foi aplicada em 15 ratos, havendo inversão na ordem de secção do nervo e do músculo. Nestes ratos, observou-sé que as PV estavam paralisadas em PPM em 12 animais, enquanto que em três, ou seja, 20% dos casos, houve paralisia em posição lateral (PL). Esperava-se que este fato ocorresse em 100%, e não em 20% somente, pois segundo a hipótese de Wagner-Grossman, o músculo cricotireóideo tem por função manter a prega vocal em posição adutora, e na sua ausência a PV deveria se deslocar lateralmente, em todos os casos. O resultado era o mesmo, ou seja, com a prega vocal em posição adutora, se, ao invés de cortar as fibras do músculo, procedíamos a denervação do mesmo, pela secção do nervo NLS.
Grupo C: utilizaram-se de sete ratos para secionarmos o NLR seguindo os movimentos da PV nas fases respiratórias. Em seis ratos a lesão do NLR ocorreu com a PV no início da abdução, e numa única oportunidade, com a prega no inicio da adução. Em todas as situações, a posição da PV foi sempre a mesma, ou seja, posição para-mediana (PPM). Quando o corte do nervo ocorreu com a PV em abdução, ou seja, em inspiração, notou-se que a PV se desloca rapidamente para PPM, como se fosse puxada.
DISCUSSÃO
Para Blitzer e colaboradores, em 19959, a lei de Semon-Rosenbach, que é. baseada na predominância da ação adutora dos músculos da PV, não é capaz de explicar o posicionamento adquirido pela PV, como da mesma forma a lei de Wagner-Grossman, que se baseia na explicação de que a posição da PV é regida pela contratura do músculo cricotireóideo. Estes autores sugerem que a reinervação variável e a sincinesia ou contração simultânea dos músculos antagonistas seriam responsáveis pelo posicionamento da PV. Woodson, em 1993 a b5, 6, também acredita que a posição assumida pelas PV nas paralisias depende da regeneração da inervação ou da inervação residual. No nosso ver, a reinervação de determinados músculos explica os casos em que ocorrem mudanças de posição da PV com o passar do tempo.
TABELA 1 - Distribuição dos animais segundo o grupo do experimento, o nervo e o músculo seccionado e o resultado obtido.
Porém, no presente estudo, a reinervação não explica os fenômenos observados, pois as posições assumidas pelas pregas vocais eram constatadas imediatamente após o trauma do nervo, não dando tempo para que a reinervação ocorresse.
Em nosso experimento, no grupo A não ocorreu mudança na posição das pregas vocais quando se realizava corte em alturas diferente do NLR ou do nervo vago. Este fato sugere que a distribuição das fibras adutoras e abdutoras- a mesma em toda extensão do nervo NLR e até mesmo do nervo vago, havendo predomínio das fibras adutoras. Porém o grupo B encontramos 20% de paralisias, em que a PV permaneceu em abdução. Este fato nos leva a afirmar que o predomínio das fibras adutoras nem sempre existe, e que a lei de Semon- Rosenbach não é aplicável nos ratos.
Para Nemiroff, em 19827, o nervo NLR no nível da borda inferior da cartilagem cricóide, Ou mais precisamente, segundo Katz, em 1986, a 0,5 cm antes da cartilagem, se divide em dois ou três ramos, denominados ramos extralaríngeos, os quais apresentam tamanhos diferenciados. Os ramos menores se dirigem posteriormente, enquanto que os maiores para a região anterior. Segundo Lang e colaboradores, em 198610, os posteriores somam-se em média 6,2 (112) ramos os quais se dirigem para o músculo cricoaritenóideo posterior, e 1,4 (1-3) ramos para o ariaritenóideo, enquanto que os anteriores totalizam cerca de 3,5 (1-6) ramos que se dirigem para o músculo cricoaritenóideo lateral e 2,06 (1-5) ramos para o músculo tireoaritenóideo. Em nosso experimento, o número de subdivisões da inervação residual não parece ter influenciado na posição final das pregas vocais nos experimentos A, B e C, pois o nervo NLR era cortado no mínimo 1 cm antes da cartilagem cricóide, Ou seja, antes dessas bifurcações.
Segundo Sanders, em 199311, à medida que o NLR caminha pelo interior da laringe, este vai dando origem a ramos para os diversos músculos. Assim, o primeiro ramo vai inervar o músculo cricoaritenóideo posterior (abdutor). Em alguns casos, existem dois ramos, um para a parte lateral e outro para medial do músculo. Estes ramos, do músculo cricoaritenóideo posterior, geralmente fazem conexões com ramos do ariaritenóideo (adutor). O próximo ramo do NLR vai para o ariaritenoideo, onde na linha média existe conexão também com ramos do NLS. Em seguida, o NLR emite ramos para o músculo cricoaritenóideo lateral (adutor) e termina no músculo tireoaritenóideo (adutor). Este fato poderia explicar porque a prega vocal ficou paralisada em PPM na maioria das vezes; corno observamos nos grupos A e B, pois O impulso nervoso demora mais para chegar ao músculo adutor. Ou seja, enquanto o abdutor está paralisado, existem adutores ainda em funcionamento, fazendo com que a PV fique em PPM. Por outro lado, verificamos gare no experimento do grupo C não houve mudanças no posicionamento da PV, ou seja, quando se procedia ao corte do NLR com a PV em posições diferentes, segundo as fases do ciclo respiratório. Este fato demonstra que a posição final da PV não depende da seqüência em que Os músculos são paralisados.
Para Exner, citado por Bertelli12, o músculo cricoaritenóideo está inerva,do pelo ramo inferior elos dois laríngeos superiores. Este detalhe anatômico poderia explicar a posição PPM ela PV, ao denervar o músculo cricotireóideo no grupo B, pois as fibras do NLS do lado oposto manteriam o tônus desse músculo responsável pela adução da PV. Porém, na maioria dos ratos do grupo 13 procedemos à secção elas fibras musculares e a PV não adquiriu a posição em abdução esperada. Sanders, em 199311, também afirma que o músculo tireoaritenóideo recebe fibras dos dois laríngeos superiores e também do NLR do mesmo lado, enquanto que o ariaritenoideo recebe fibras dos dois NLS e dos dois NLR. Este fato também não explica o posicionamento das PV em nosso experimento, uma vez que na maioria dos ratos o experimento era realizado de um lado e depois do outro. Ora, quando se cortava os nervos NLR e NLS, de um lado, a PV ficaria em PPM, às custas da contração desses músculos devido aos impulsos enviados pelo nervo do lado oposto. Porém, ao se cortar os nervos do outro lado, a PV deveria ficar em PL, o que não ocorreu.
Tschiassny, em 19572, afirmava que a secção cio cricotireóideo ou denervação do músculo pode não ser capaz de promover a melhora da dispnéia nos pacientes com paralisia das PV em PPM, porque o lado paralisado tende a ser aspirado pela pressão negativa intralaríngea no momento da inspiração. Assim, segundo o autor, o efeito de Bernoulli seria responsável por manter a PV em PPM. Pelos resultados de nosso estudo, e corri base em nossa experiência clínica, não acreditamos neste fato, pois haveria movimento da PV ao se, sugada pela pressão negativa. A pequena movimentação da PV paralisada, observada na inspiração, é decorrente da contratura cios músculos extrínsecos da laringe, ou seja, aqueles do pescoço.
Segundo Koufman e colaboradores, em 199513, além do músculo cricotireóideo não influenciar na posição final elas PV nas paralisias, também a contração cio músculo tireoaritenóideo parece não influenciar na posição final da prega vocal. Concordamos com eles em que fatores até agora desconhecidos determinam a posição da PV nas paralisias da laringe; porém, salientamos que ao se cortar o NLR durante a inspiração, ou seja, em abdução a PV adquire um movimento brusco em direção à linha mediana, como se fosse puxada por um sistema elástico.
CONCLUSÕES
Com base nas observações desse trabalho, pode se verificar que, em ratos:
- o músculo cricotireóideo não exerceu influência na determinação da posição final da PV;
- a posição da PV não dependia do último músculo intrínseco contraído;
- a PV se desloca bruscamente para PPM quando o corte do NLR ocorre na inspiração;
- em 20%dos ratos a PV ficou em PL nas paralisias dos NLR e NLS.
REFERÊNCIAS BIBLIOGRÁFICAS
1. BLITZER, A.; JAHN, A. F. & KEIDAR, A. - Semon s law revisited: na electromyography analysis of laryngeal synkinesis. Ann. Otol. Rhinol. Laryngol., 105: 764- 69, 1995
2. EXNER - Citado por Antonio de Paula Bertelli in Cancer da Laringe. Ed. Manole, São Paulo, 1980.
3. FREEDMAN, L. M. -The role of the cricothyroid muscle in tension of the vocal cord. Arch. Otolaryng., 62:347- 53, 1955
4. KATZ, A. D. - Extralaryngeal division of the recurrent laryngeal nerve. Report on 400 patients and the 721 nerves measured. Am. J. Surgery, 152: 407-10, 1986
5. KOUFMAN, J. A.; WALKER, F. O. & JOHARJI, G. M. - The cricothyroid muscle does not influence vocal flod position in laryngeal paralysis. Laryngoscope 105: 368- 72, 1995.
6. LANG, J.; NACHBAUR, S. & FISCHER, K. - Laryngeal nerves, branches in the interior of the larynx. Gegenbaurs Morphologisches Jahrbuch, 132: 723-36, 1986
7. MURAKAMI, Y. & KIRCHNER, J. A. - Vocal cord abduction by regenerated recurrent laryngeal nerve. Na experimental study in the dog. Arch. Otolaryng., 94: 64- 68, 1971
8. NEMIROFF, P. M. & KATZ, A. D. - Extralaryngeal divisions of the recurrent laryngeal nerve. Surgery and clinical significance. Am. J. Surgery, 144: 466-9, 1982
9. SANDERS, I.; WU, B. L.; UM, L.; LI, Y. & BILLER, H. F. -The inervation of the larynx. Arch. Otolaryngol. Head Neck Surg., 119: 934- 39, 1993
10. SUZUKI, M.; KIRCHNER, J. Á. & MIJRAKAMI, Y. - The cricothyroid as -a respiratory muscle: its characteristics in bilateral recurrent laryngeal paralisys. Ann. Otol., 79: 97683, 1970
11. TSCHIASSNY, K. - Therapeutically induced paralysis of the cricothiroid muscle or its removal in paralysic laryngeal stenosis. Arch. Otolaryng., 65: 133- 42, 1957
12. WOODSON, G. E. - Configuration of the glottis in laryngeal paralysis. I: clinical study. Laryngoscope, 103:1227- 33, 1993 (a).
13. WOODSON, G. E. - Configuration of the glottis in laryngeal para-lysis.II: animal experiments. Laryngoscope, 103: 123541, 1993 (b).
* Chefe do Serviço de Cirurgia de Cabeça e Pescoço.
** Professor Assistente.
*** Médico Residente. Trabalho realizado no Serviço de Cirurgia de Cabeça e Pescoço do Hospital das Clínicas e Departamento de Cirurgia, Ortopedia e Traumatologia da Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo.
Trabalho apresentado no XVII Congresso Brasileiro de Cirurgia de Cabeça e Pescoço, no período 3 à 6 ele setembro ele 1999, em Belo Horizonte/ MG.
Endereço para correspondência: Professor Dr. Rui Celso Martins Mamede-Hospital das Clínicas ela Faculdade ele Medicina de Ribeirão Preto da Universidade de São Paulo Campus Universitário - Monte Alegre - 14048-900 Ribeirão Preto/ SP - Telefone/fax: (Oxx16) 602-2353 - E-mail:- rcmmamed@rgm.fmrp-usp.br
Artigo recebido em 8 de novembro de 1999. Artigo aceito em Vele dezembro de 1999.
Imprimir: ![]()