

Ano: 1983 Vol. 49 Ed. 2 - Abril - Junho - (2º)
Seção: Artigos Originais
Páginas: 17 a 28
FISIOLOGIA DO SISTEMA VESTIBULAR
Autor(es): JOSÉ ANTONIO APPARECIDO DE OLIVEIRA 1
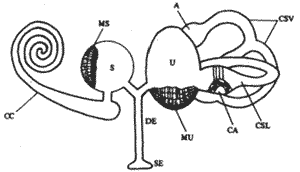
![]()
CC - Canal Cocka
MS - Máculã Swvlar
S - Sáculo
DE - Douto Endolinfático
SE - Saco Endolinfítico
CA - Crista Ampolar
U - Utrímo
CSV - anais Semicirculares Vertìcais
MU - Mácula Utricular
A- Ampolá
ClS - Canais semicirlares Laterais
fig. 1 - Const~ do sistema vestibular.
Os receptores de posição e movimento da cabeça estão localizados no sistema vestibular. 0 ouvido interno apresenta no osso temporal duas partes que constituem o labirinto. O labirinto anterior é a cóclea e o labirinto posterior é o sistema vestibular.
Constituição do sistema vestibular - (Fig. 1)
É uma estrutura óssea no osso temporal que contém no seu interior estruturas membranosas. As estruturas membranosas são: o século, o utrículo e três canais semicirculares. Entre as paredes das cavidades ósseas e as paredes das estruturas membranosas existe um líquido, a perilinfa, e dentro das estruturas membranosas temos a endolinfa.
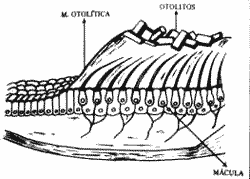
Estrutura do sistema otolítico - (Fig. 2)
Os órgãos otolíticos estão localizados nas vesículas, utrículo e Báculo. Cada órgão otolítico é formado por uma membrana basal que sustenta as células chiadas. Sobre estas células sensoriais há uma estrutura acessória fornada por uma massa gelatinosa onde estão mergulhados os cílios da células sensitivas. A massa gelatinosa contém muitos cristais de carbonato de cálcio, de pequenas dimensões formando assim a membrara otolítica. Estes cristais são mais densos do que o material gelatinoso e são chamados otolìtos.
O conjunto das células ciliadas e células de sustentação que constitui a estrutura receptora é chamado mácula. Assim existe a mácula sacular com sua membrana otolítica, no século e a mácula utricular com sua membrana otolítica, no utrículo. A mácula secular apresenta-se em posição verticalizada e a utricular horizontalizada, quando o corpo se encontra em posição ereta. As células sensoriais das máculas apresentam dois tipos de cílios: os Kinocilios, em número de um para cada célula, sendo maior e situado lateralmente e com ultra-estrutura de um cílio motor de protozoário, os estereocilios são mais finos, curtos e numerosos colocados lateralmente ao Kinocílio em comprimentos decrescentes, quanto mais distantes do mesmo. Em cada mácula diferentes grupos de células ciliadas estão orientadas em direções diferentes com seus Kinocílios em direções opostas. Nos pólos inferiores das células sensoriais encontramos células nervosas fazendo sinapse. As células sensitivas do sistema vestibular são células epiteliais modificadas constituindo receptores secundários.
Fig. 2 - Estrutura do sistema otoiltico.
Estímulo
Os estímulos para os receptores otolíticos são a aceleração gravitacional e outros tipos de acelerações lineares. Movimentos num elevador, parada brusca de um carro, por exemplo.
Mecanismo de estimulação-transdução
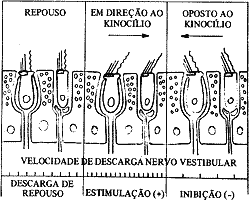
Quando a cabeça está em posição vertical a mácula utricular estará em posição horizontal e em repouso, não havendo estimulação efetiva pois os cílios estarão verticalizados sem forças tangenciais atuando sobre eles. Neste caso ocorre uma descarga de impulsos espontânea, de repouso nos axônios, que se ligam às células sensitivas. Na mácula sacular devido sua posição verticalizada, estará ocorrendo uma estimulação efetiva. Isto porque os otolitos atraídos pela força gravitacional provocam deslocamento das membranas otolíticos em relação ao epitélio sensorial da mácula. Assim, os estereocílios se deslocam em relação aos Kinocílios devido às forças tangenciais a que são submetidos. Esta inclinação dos cílios altera a permeabilidade da membrana da célula sensorial, provocando fluxo de íons sódio para o interior da célula, diminuição do potencial de membrana negativa e conseqüente despolarização com formação do potencial gerador. Este potencial gerador dependerá da intensidade do estímulo e quanto maior seu valor determinará o aparecimento de uma freqüência de descarga de impulsos maior nos axônios das células nervosas da mácula. Como as células sensoriais são agrupadas de modo diferente, com seus cílios em posições opostas na mácula, conforme a posição da cabeça existirão células estimuladas positivamente (deslocamento dos estereocílios em direção ao Kinocílio) e células estimuladas negativamente (deslocamento do Kinocílio em relação aos estereocílios). No primeiro caso a freqüência de descarga de impulsos nos axônios será maior que a freqüência de repouso e no segundo caso a freqüência será menor que a de repouso. Quando a cabeça forma um ângulo com a vertical inclinando-se, haverá deslocamento dos órgãos otolíticos e, devido a gravidade, formação de forças tangenciais em diferentes máculas e diferentes áreas receptoras em cada mácula.
Deste modo todas as posições da cabeça podem ser captadas pelos axônios das máculas que enviam estas informações a várias localizações cerebrais. Estas informações provocam movimentos reflexos que permitem adaptação da posição do tronco e dos membros com relação à posição da cabeça, mantendo assim o equilíbrio. O sistema otolítico associado a proprioceptores dos músculos, articulações, receptores cutâneos e visuais permitem a regulação do equilíbrio estático do corpo.
Nota - O sáculo nos vertebrados inferiores tem função de equilíbrio e detecta sons graves. No homem estas funções são discutidas havendo controvérsias.
Adaptação
Os órgãos otolíticos não detectam modificações na posição da cabeça, eles assinalam a posição atual da cabeça, não apresentando praticamente o fenômeno de adaptação, como a maioria dos outros sistemas sensoriais. Se alguma adaptação ocorrer pode ser devido a modificações neurais.
Estrutura dos canais semicirculares
São três canais cheios de endolinfa e fechados, situados no osso temporal em posições perpendiculares entre si.
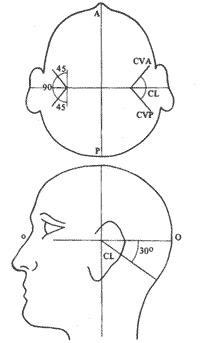
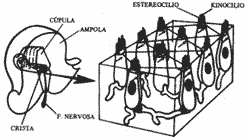
Numa das extremidades de cada canal está uma dilatação, a ampola. De cada lado do crânio temos três canais semicirculares: o horizontal ou lateral, o vertical anterior e o vertical posterior. O canal horizontal na realidade não ocupa o plano horizontal quando a cabeça está em posição normal, pois apresenta em relação ao plano horizontal 300 com seu bordo anterior, a ampola discretamente desviada para cima (Fig. 3). Todo o sistema descrito com o sáculo, o utrículo e os três canais semicirculares constituem com a cóclea o labirinto membranoso cheio de endolinfa que fica localizado em cavidades ósseas correspondentes que constituem o labirinto ósseo. Na ampola do canal semicircular temos as células receptoras ciciadas que juntamente com as células de sustentação formam a crista ampolar. Como estrutura acessória temos a cúpula gelatinosa que faz saliência no líquido e enche o interior da ampola, sendo seu peso específico igual ao da endolinfa, por isso não se desloca durante aceleração linear. Como a cúpula preenche a ampola, a endolinfa não flui ao redor dela. Os estereocílios e Kinocílios das células da crista estão mergulhados na cúpula. Dos pólos inferiores das células sensoriais que são receptores secundários partem fibras nervosas que vão formar o nervo vestibular. (Fig. 4).
Fig. 3 - Localização dos canais semicirculares.
Fig. 4 - Estrutura do receptor ampolar.
Estímulos
Movimentos da cabeça em qualquer direção afetam pelo menos um canal semicircular, assim os canais percebem toda rotação da mesma. Deste modo as cristas são estimuladas por acelerações angulares de no mínimo 1 grau por segundo, daí ocorrerão reflexos estato-sinéticos para regulação da postura.
Mecanismo de estimulação - transdução
Vamos considerar o canal horizontal
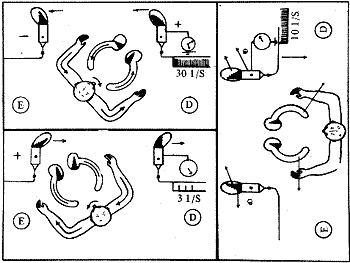
Quando o crânio é movimentado no plano horizontal imprimindo-se a ele uma aceleração angular, esta aceleração afeta a cúpula devido ao deslocamento da endolinfa pela sua inércia. O deslocamento da endolinfa é oposto ao da rotação da cabeça, constituindo uma corrente de endolinfa em direção à ampola (empulópeta) isto no canal horizontal do lado da rotação. No canal oposto a endolinfa deslocar-se-á em direção oposta à ampola (corrente ampulófuga).
Quando por exemplo giramos a cabeça para a direita a corrente ampulópeta será no canal horizontal direito e esta determinará um deslocamento da cúpula em direção ao utrículo, o que acarretará um deslocamento dos estereocílios em direção ao Kinocílio, pois este está colocado em direção ao utrículo nos canais horizontais, sendo sua posição contrária nos canais verticais. Este padrão de inclinação dos cílios da crista ampolar direita, por forças tangenciais, provocará alteração da permeabilidade da membrana das células sensoriais e diminuição do potencial negativo de membrana, pela entrada na célula de íons sódio, ocorrendo conseqüentemente uma despolarização. Deste modo ocorre o fenômeno de transdução aparecendo o potencial gerador e havendo uma estimulação positiva (efetiva). A freqüência de descarga de equilíbrio que existia nos neurónios aumenta devido esta estimulação, e o sistema nervoso central é informado que a cabeça girou para o lado direito. (Fig. 5)
Como do lado esquerdo a corrente foi ampulófuga, a cúpula se inclina em direção oposta ao utrículo e os Kinocílios deslocam-se em direção aos estereocílios, o que acarreta uma estimulação negativa (inefetiva), determinando uma hiperpolarização nas células receptoras e uma freqüência de descarga menor que a de repouso nos neurônios vestibulares. O sistema nervoso central, quando a cabeça gira para a direita, recebe freqüência de descarga maior dos receptores vestibulares deste lado e menor do lado oposto, concluindo que realmente a cabeça girou para a direita.
Quando o movimento é mantido, a velocidade da endolinfa é a mesma que a das estruturas dos canais, não havendo estimulação. Ao parar bruscamente o movimento que realizamos para a direita a corrente ampulópeta será do lado esquerdo com estimulação positiva deste lado e do lado direito a corrente será ampulófuga havendo estimulação negativa.
DESLOCAMENTO DOS CÍLIOS
Fig. 5 - Mecanismo de transdução nas células ciliadas da crista ampolar.
Fig. 6 - Mecanismo de estimulação do receptor ampolar com rotação da cabeça para a direita e esquerda.
Para compreender estes mecanismos facilmente podemos usar um modelo. Imagine nossos braços curvados e colocados horizontalmente como os dois canais semicirculares horizontais. As nossas mãos fechadas corresponderiam às ampolas onde estão as cristas. Nossos braços seriam canais cheios de líquido.
Ao deslocarmos a cabeça para a direita e os braços conjuntamente fica fácil imaginar que pela inércia o líquido do braço direito deslocará em direção à mão (estímulo positivo) e no braço esquerdo o líquido deslocará em sentido contrário à mão (estímulo negativo) (Fig. 6).
Adaptação
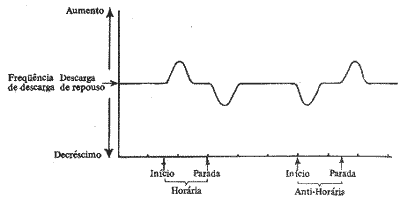
Podemos registrar os potenciais dos axônios relacionados com as cristas dos canais semicirculares. Com a cabeça em repouso existe uma freqüência de descarga espontânea de equilíbrio que é baixa. Se o animal for colocado num aparelho rotatório, quando a rotação inicia, a freqüência de descarga aumenta, para logo em seguida, durante a rotação, voltar a descarga baixa de repouso, havendo assim uma adaptação, isto ocorrendo, porque durante a rotação em velocidade constante a endolinfa tem a mesma velocidade que o canal semicircular, não havendo estimulação e deslocamento da cúpula. Quando a rotação cessa, aqueles axônios que tinham aumentado a freqüência de descarga, agora diminuem esta freqüência até um nível menor que a freqüência de repouso. Estes fenômenos elétricos estão ocorrendo nos axônios dos receptores do lado da rotação. Ao lado oposto, as variações das freqüências de descarga serão exatamente opostas. Como vemos, estes axônios assinalam mudanças nos movimentos da cabeça. Podemos colocar em gráfico o que acontece com a freqüência de descarga dos axônios dos receptores dos canais semicirculares, quando ocorre uma rotação em sentido horário e em seguida anti-horário (Fig. 7). Nós podemos dizer que a maioria dos axônios assinalam a direção da mudança de movimento.
Fig. 7 - Grafico da variação de freqüência de descarga na rotação horária e anti-horária.
Papel dos canais semicirculares no equilíbrio
Como vimos, os canais semicirculares detectam quando começamos a girar a cabeça numa direção ou quando paramos a rotação; pode-se concluir que a função dos canais não é manter um equilíbrio estático quando submetidos a forças centrífugas. A perda das funções destes receptores determina desequilíbrio quando realizamos movimentos rápidos com a cabeça.
Estas características de funcionamento permitem uma função preditiva dos canais, pois podem determinar que uma pessoa está começando a virar a cabeça. Esta informação informa ao sistema nervoso central que a pessoa estará desequilibrada dentro em pouco, daí uma necessidade de correção. Há uma predição que haverá um desequilíbrio antes que ele ocorra, permitindo que o sistema nervoso faça os ajustamentos adequados.
Vias vestibulares
As fibras nervosas aferentes que provêm das cristas ampolares ou das máculas otolíticas atingem a cavidade craniana pelo meato acústico interno, onde se encontram os corpos celulares bipolares formando o gânglio vestibular ou de Scarpa. Estas fibras vão formar a porção vestibular do oitavo nervo craniano que unindo-se à porção coclear dirigem-se aos núcleos bulbares seguindo o mesmo trajeto. Este neurônio representa o primeiro neurônio vestibular.
Do meato auditivo interno seguem as fibras até os núcleos vestibulares bilaterais localizados no bulbo próximo à linha média em relação com o assoalho do IV ventrículo inferiormente. São os núcleos vestibulares bulbares de Deiters, Schwalbe, Bechterev, Roller, Brodal. Nestes núcleos encontramos as células do segundo neurônio. O segundo neurônio vestibular é um neurônio de associação com os núcleos oculomotores, medulares, neurovegetativos e constitui o "cierre" do arco reflexo vestíbulo efetor, muscular ou neurovegetativo. (Fig. 8)
Fig. 8 - Vias vestibulares e principais conexões.
Conexões vestibulares - (Fig. 8)
Conexões oculomotores. (Vias vestíbulo-oculomotores.) Formação reticular
A projeção do estímulo vestibular aos núcleos oculomotores produz a coordenação dos olhos com a posição da cabeça e corpo no espaço. O aumento da freqüência de descarga de impulsos originado, por exemplo, nas células receptoras da crista ampolar do canal horizontal é transmitido aos núcleos oculomotores contralaterais pois as fibras que saem dos núcleos vestibulares cruzam a linha média antes de alcançar o núcleo do nervo craniano motor ocular comum (III) e do nervo abducente (VI). 0 primeiro inerva o músculo reto medial do lado oposto ao núcleo e o segundo o reto lateral do mesmo lado.
Conexões medulares (vias vestíbulo-espinais)
Estas conexões realizam-se em grande parte através do núcleo de Deiters que recebe também fibras do cerebelo de modo que o sistema vestibular e o cerebelo juntos possam regular o tono muscular do pescoço, troncos e extremidades de ambos os lados. O aumento da freqüência de descarga, por exemplo, nas células receptoras da crista do canal horizontal determina a transmissão do impulso (potencial de ação) até os núcleos dos nervos espinais do lado oposto principalmente, o que determina o aumento do tono em toda musculatura contralateral do lado estimulado, musculatura extensora do pescoço, tronco, extremidades.
Conexões cerebelosas - Lóbulo nódulo-flocular
O cerebelo recebe o estímulo propriorreceptivo vestibular, tanto do primeiro neurônio como dos núcleos de Betcherew e Deiters. Estes impulsos provenientes do sistema vestibular associam-se aos impulsos propriorreceptivos da sensibilidade profunda e estímulos estéreo e interoceptivos do organismo. Deste modo, pela via cerebelo-vestíbulo-núcleo de Deiters espinal, mantém o tono muscular adequado para conservar o equilíbrio e os movimentos.
Conexões neurovegetativas - São múltiplas
a) com o núcleo vegetativo hipotalâmico
b) com a substância reticular bulbar e mesencefálica
c) com o pneumogástrico (vago)
Conexões corticais - Córtex parietal
O sistema vestibular é um órgão sensorial e para tanto deve ter um componente consciente. A mais importante via parece ser a que envia ao cerebelo, região talâmica e cortical, o impulso proveniente de estímulos proprioceptivos vestibulares e de sensibilidade profunda.
Estes estímulos de sensibilidade profunda devidos à contração muscular, estiramento de tendões e posição das articulações informam ao córtex as posições dos segmentos do corpo no espaço. A via vestibular informa a posição da cabeça estática ou dinâmica no espaço devido estímulos provenientes do sistema vestibular.
O conjunto destes estímulos dá a sensação de equilíbrio.
Respostas reflexas
Músculos do olho - Nistagmo
O aumento do tono dos músculos do olho, reto lateral do lado oposto ao estimulado e reto medial do olho do lado estimulado provoca um desequilíbrio de tonos em relação aos músculos antagonistas e os dois olhos são desviados conjugadamente para o lado oposto ao estímulo.
Este desvio de origem vestibular corresponde à fase lenta do nistagmo e a um movimento compensatório dos olhos para reter o campo do último olhar, oposto ao movimento da cabeça, pois como vimos, se a cabeça gira para a direita, horizontalmente, por exemplo, este desvio é para a esquerda. Este é um mecanismo protetor, retendo o sistema nervoso, a orientação do meio ambiente. A mesma ação do exemplo citado pode ser obtida estimulando-se com, água quente o canal horizontal direito de um paciente em decúbito dorsal com a cabeça elevada de 300 em relação ao plano horizontal.
A volta do olho em sentido contrário constitui a fase rápida do nistagmo. A direção do nistagmo é dada por este componente rápido. Para seu aparecimento a substância reticular é indispensável.
Aplicando estímulos rotatórios e calóricos respectivamente a velocidade dos olhos durante a componente lenta é proporcional ao estímulo aplicado no sistema vestibular.
Músculos espinais - Desvios
O aumento do tonos dos músculos dos segmentos do corpo do lado oposto ao estímulo provoca desequilíbrio
em relação ao tonos dos músculos do lado oposto; isto pode determinar a queda do indivíduo para o lado contrário ao estimulado.
Núcleos vagais - Manifestações neurovegetativas
A freqüência de descarga dos impulsos nervosos provenientes dos receptores vestibulares pode determinar, devido aos núcleos vagais, reações neurovegetativas como náusea, vômito, queda de pressão, sudorese, diarréia.
Teoria do equilíbrio
Em condições normais a estimulação (rotação) provoca uma modificação da descarga de impulsos de repouso dos dois lados. De um lado o potencial de repouso é modulado num sentido e do outro em sentido contrário. A diferença de modulação é a mesma dos dois lados em sentidos contrários. Deste modo os dois lados do cérebro são informados de um modo igual mas oposto. Esta polaridade é importante para se compreender o sistema.
Esta polaridade normalmente é recebida pelas estruturas com as quais o sistema vestibular mantém conexões.
a) o córtex interpreta a modificação como um movimento na direção e velocidade específica.
b) os núcleos dos músculos oculares movem os olhos compensatoriamente, para reter o campo do último olhar, oposto ao movimento da cabeça, como um mecanismo protetor retendo a orientação do meio ambiente.
c) as células do corno anterior da medula espinal ajustam o tonos dos músculos do tronco e membros.
d) o cerebelo ajusta o tono para a nova situação.
Estes processos são em parte instintivos e parcialmente aprendidos.
O córtex aprende relacionar o grau de inclinação da cúpula em resposta a movimentos repentinos da cabeça, com o grau específico de alteração do meio ambiente por equiparação; os olhos informam que o corpo moveu, os proprioceptores também, modalidades tácteis informam ao cérebro quando o organismo toca o solo. Durante anos o cérebro aprende a combinar as diferentes modalidades com o sistema vestibular, originando-se, então, a sensação vestibular. Sendo um computador perfeito, na infância já integrou estas sensações.
Podemos dizer que a finalidade dos dois sistemas vestibulares é a de informar o organismo de movimentos e posição da cabeça para o ajuste do corpo a estas novas posições.
Comparando este esquema de funcionamento e a estrutura do sistema vestibular podemos interpretar à luz destes conhecimentos os sintomas provocados por doenças vestibulares, sintomas estes narrados pelo paciente e os sinais que poderão ser detectados pelos testes labirínticos e eletronistagmografra.
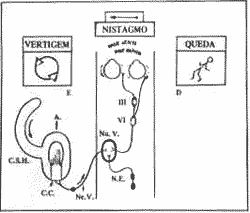
Importância clínica do nistagmo Crise Labiríntica (Fig. 8)
Quando há um distúrbio num labirinto, esquerdo por exemplo, resultando em diminuição repentina da freqüência de descarga, ele é incapaz de contrabalançar o lado normal com uma descarga oposta e igual.
O cérebro capaz de interpretar a luz de respostas aprendidas passadas distribui uma falsa corrente de impulsos a seus efetores. Deste modo surge a vertigem; o córtex é informado que o organismo está em um estado constante de movimento.
Os olhos procuram repetítivamente reter o campo do último olhar (nistagmo), os músculos dos membros preparam-se para encontrar uma situação que é irreal (desequilíbrio e queda) e o intestino pára e começa a peristalse reversa (náusea e vômito).
O lado envolvido (doente, hipofuncionante como na doença de Ménière) não é capaz de comunicar ao cérebro sua informação de repouso. Deste modo os dois sistemas vestibulares estão descarregando em repouso em uma intensidade desigual o que tem para o cérebro um significado específico. As seqüelas deste desequilíbrio são manifestações de uma hiperfunção relativa do lado intacto; então como resultado aparecem reflexos vestibulares prolongados e incontrolados.
Fisiologia da compensação
O estado de desequilíbrio não poderia durar muito tempo, o organismo eventualmente morreria através da desidratação e desequilíbrio fluido e eletrolítico. Deste modo o organismo tenta equilibrar a situação. A restauração do equilíbrio entre os dois centros inicia o controle dos reflexos patológicos.
Esta restauração pode ocorrer por supressão central do lado intacto por invocação de tratos inibitórios no sistema nervoso central, possivelmente do cerebelo.
Também pode haver geração de uma nova atividade elétrica no sistema, com descarga menor, para equilibrar lado normal, hipercinético em relação ao lado oposto ela condição patológica. Após dias ou semanas da lesão o órgão terminal esta nova atividade de repouso do úcleo denervado determina o desaparecimento dos intomas e a inibição cerebear vai desaparecendo agarosamente. Não se sabe o que estimula a geração de nova atividade de repouso no núcleo denervado, mas sabemos que é necessário o desequilíbrio provocando uma espécie de vácuo, que estimula a necessidade do seu aparecimento.
Além do mais o organismo tenta fazer o órgão doente voltar à normalidade.
Esta restauração torna a situação tolerável para o organismo desde que não haja estimulação do lado bom o que acentua o desequilíbrio. Daí, durante a crise o paciente se mantém bem sem movimento da cabeça.
Isto acontece nas várias doenças vestibulares como por exemplo na doença de Ménière, labirintite supurativa etc.
A rapidez da recuperação dependerá da idade do paciente, eficiência do bloqueio neuronal, eficiência do suprimento vascular do sistema nervoso central. Este mecanismo de compensação tem uma implicação clínica muito importante; se os sintomas persistem continuamente por mais de duas ou três semanas a causa não é vestibular. Este fato associado a crise significante sem nistagmo labiríntico espontâneo nos indicam que o distúrbio não é vestibular.
Coordenação do equilíbrio pelo cerebelo
As informações da posição estática e dinâmica da cabeça são enviadas entre outras regiões do sistema nervoso central ao cerebelo. Estes impulsos chegam aos lóbulos floculonodulares, regiões importantes para a integração adequada dos impulsos do equilíbrio, especialmente alterações da direção do movimento.
A extirpação daquelas regiões cerebelares determina sintomas de desequilíbrio que são os mesmos apresentados na destruição dos canais semicirculares. O cerebelo sabe com antecipação que rotações do corpo provocarão desequilíbrio, isto permite uma coordenação da função muscular, ocorrendo os ajustes corretivos posturais necessários nas atividades motoras do córtex cerebral, para a manutenção do equilíbrio e postura.
O cerebelo recebe informações também dos fusos neuromusculares, órgãos tendinosos de Golgi, receptores tácteis da pele e articulações, que informam o estado atual da contração muscular, tensão dos tendões, posições das partes do corpo. Assim o cerebelo mantém-se informado sobre a posição do corpo parado ou em cada instante. Também o cerebelo recebe informações do córtex motor sobre os estímulos motores que estão atuando nos músculos e articulações. Deste modo o cerebelo recebe as informações referentes à posição do corpo, pelos órgãos sensitivos já citados, recebe as informações dos estímulos corticais aos músculos do corpo, podendo então comparar as duas situações e enviar ao córtex comandos de reajustes para manutenção do equilíbrio.
Provas de estimulação vestibular
Podemos submeter um indivíduo ou animal a estimulação vestibular através de várias provas usadas em clínica e em pesquisa e analisar suas reações com relação a vertigem nistagmo e desvio.
1. Prova rotatória
Devemos analisar as reações reflexas, após a parada do movimento rotatório, estando a cabeça do indivíduo inclinada 30o para frente, horizontalizando assim os canais horizontais.
a) Pós-rotação para a direita
Canal horizontal esquerdo - corrente ampulópeta, despolarização, estimulação efetiva do receptor (+)
Canal horizontal direito - corrente ampulófuga, hiperpolarização, estimulação inefetiva do receptor (-).
Reações:
Vertigem para a esquerda
Nistagmo para a esquerda
b) Pós-rotação para a esquerda
Canal horizontal direito - corrente ampulópeta, despolarização, estimulação efetiva do receptor (+).
Canal horizontal esquerdo - corrente ampulófuga, hiperpolarização, estimulação inefetiva (-).
Reações:
Vertigem para a direita
Nistagmo para a direita
Desvio para a esquerda
2. Prova calórica
Podemos estimular o sistema vestibular irrigando-se os ouvidos com água quente ou fria, desde que a cabeça do indivíduo fique com o canal semicircular verticalizado com a ampola para cima. Isto pode ser feito com o indivíduo deitado com a cabeça levantada em 30º com relação ao plano horizontal ou num indivíduo sentado com a cabeça inclinada 60º para trás em relação ao mesmo plano. A ampola com o receptor nos dois casos ficará em posição superior. Esta estimulação artificial com água quente ou fria baseia-se no fato de o calor propagado ao labirinto provocar correntes de convecção na endolinfa, fazendo esta deslocar-se para a ampola (água quente) e contra a ampola (água fria). Analisando as reações teremos:
Fig. 9 - Registro elétrico do nistagmo.
a) Água quente no ouvido direito (44ºC) - corrente ampulópeta, despolarização, estimulação efetiva do receptor (+).
Reações:
Vertigem para a direita
Nistagmo para a direita
Desvio para a esquerda
b) Água quente no ouvido esquerdo (44ºC) - corrente ampulópeta, despolarização, estimulação efetiva (+).
Reações:
Vertigem para a esquerda
Nistagnio para a esquerda
Desvio para a direita
c) Água fria no ouvido direito (30ºC) - corrente ampulófuga, hiperpolarização, estimulação, inefetiva no receptor ().
Reações:
Vertigem para a esquerda
Nistagmo para a esquerda
Desvio para a direita
d) Água fria no ouvido esquerdo (30°C) - corrente ampulófuga, hiperpolarização, estimulação inefetiva (-)
Reações:
Vertigem para a direita
Nistagino para a direita
Desvio para a esquerda
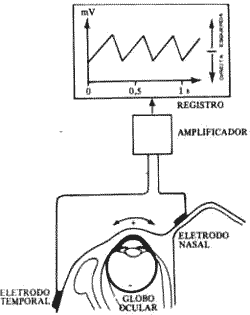
Elestronistagmografia (Fig. 9)
Como a córnea é eletricamente positiva em relação à retina, se colocarmos um eletrodo em cada lado externo dos olhos de um na fronte podemos registrar as variações de potencial, que ocorrem durante o nistagmo, com um aparelho eletronistagmógrafo e o registro será a eletrnistagmografia. Este exame é usado no homem para estudar o nistagmo normal e suas variações nas doenças labirinticas. Em animais podemos usar o método, para avaliar o efeito de drogas vestibulotóxicas como os antibióticos aminoglicosidicos, estreptomicina e gentamicina, por exemplo.
Estes estudos, que têm sido feitos em cobaias principalmente, podem ser complementados por estudos histológicos de preparação de surperficie ao microcóspio de contraste de fases, assim serão detectadas as lesões de epitélio sensorial vestibular.
REFERÊNCIAS BIBLIOGRÁFICAS
ALONSO, C.G. Anatomia y Fisiologia dei labirinto posterior. Madrid: Editorial Paz Montalvo, 1966.
BANCHIERI, G.C. Le sindromi vertiginose nell'anziano. Giorn. Geront. 22: 203-213, 1974.
BARTALENA, G. I disturbi vestibulare nella senescenza. Giorn. Geront. 25: 99-106, 1977. BRODAL, A.
POMPERANO, O. Basic aspects of central vestibular mechannisms. Amsterdam: Elsevier Publishing, 1972.
BRUNÁS, R.L., MARELLI, E.F. Sistema vestibular. Rosário, Argentina: Llordén Soc. Resp. Ltdr. , 7.
CASE, J. Sensory Mechanisms. London: Collier-McMWan Limited, 1967.
DUFOUR, A., MIRA, E., PIGNATARO, O. Vestibologia, Milano-Italia: Aldo Gorzonti Editore, 1980.
GOLDBERG, J.M. and FERNANDEZ, C. Vestibular mechanisms. Ann. Rev. Physiol., 37:129,1975.
HOOD, J.D. Vestibular mechanisms in health and disease. London: Academic Press, 1978.
LOWESTEIN, O. The senses. Inglaterra: Chaucer Press Ltda., 1966.
LUDEL, J. Introduction to Sensory processes. San Francisco: W.H. Freeman and Company, 1978.
MELLON, D.J. The physiology of Sense Organs. London: Oliver and Boyd Ltd.,1968.
OLIVEIRA, J.A.A. Fisiopatologia do sistema vestibular. Revista Brasileira de Otorrinolaringologia, 40: 64, 1974.
OLIVEIRA, J.A.A. Uma técnica para o estudo da função vestibular na cobaia. Revista Brasileira de Otorrinolaringologia, 43:264,1977.
SCHIMIDT, R.F. Fundamentais of Sensory physiology. Berlin: Springer Verlag, 1978.
WOLFSON, R.J. The Vestibular System and its Diseases. Phìladelphia: University of Pennsylvania Press, 1966.
1 - Professor adjunto - Laboratório de Bioacustia Professor Ricardo F. Marseillan - Faculdade de Medicina - Ribeirão Preto - USP. Laboratório de Biocomunicapão - Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto - USP. Departamento de Of talmo-Otorrinolaringologia - Faculdade de
Medicina de Ribeirão Preto - USP.
Imprimir: ![]()
All rights reserved - 1933 /
2026
© - Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico Facial