

Ano: 1975 Vol. 41 Ed. 2 - Maio - Agosto - (8º)
Seção: Trabalhos Originais
Páginas: 171 a 179
OLFATO - ATUALIZAÇÃO (1.ª Parte)
Autor(es):
Ary Guilherme Ferreira
Alvaro Acar
Resumo:
O trabalho, dividido em 3 capítulos histologia da mucosa olfativa, anatomia das vias e centros e eletrofisiologia relata os conceitos mais modernos sobre o assunto.
![]()
O estudo do sentido do olfato não tem sido olhado com o interesse e importância devidos. Nos últimos anos, graças à ultra-microscopia eletrônica, aos progressos da eletrofisiologia, da cromatografia e espectrofotometria, os pesquisadores estão penetrando e desvendando os segredos do intrincado universo olfatório. Estes conhecimentos consubstanciaram-se com os trabalhos de Mc Leod, Ottoson, Amoore, Fortunato e Nicolini, Le Magnen, Graziadei, e outras. Atualmente, já sabemos como se processa, onde se realiza e como o estímulo olfatório é conduzido aos centros corticais. A olfação efetua-se através de um sistema que vai da vesícula olfatória, localizada na mucosa nasal, até a córtex cerebral. Devido à sua unidade histológica podemos afirmar que se trata mais de um sistema do que de um órgão ou aparelho.
Nessa primeira etapa desenvolveremos os capítulos referentes à histologia da mucosa olfatória, à anatomia das vias e centros e à eletrofisiologia.
ESTUDO DA MUCOSA OLFATÓRIA - A mucosa olfatória está situada na parte póstero-superior das fossas nasais, não existindo uma linha limitativa precisa entre a mucosa olfatória e a mucosa respiratória. Esta mucosa apresenta uma coloração amarelo-escuro motivada pela presença de um cromolipídio característico da função olfatória. Moncrief dá muita importância a esse pigmento no mecanismo da olfação. Assim, os porcos brancos da Virginia e os carneiros brancos de Trentino são preteridos na criação por comerem plantas venenosas devido ao seu pobre sentido do olfato. Os albinos são conhecidamente anósmicos congênitos, sendo a ligação albinismo-hiposmia uma conseqüência da escassez em pigmentos de sua área olfatória.
Há dois tipos de pigmentos amarelos na mucosa: os carotenóides (carotenóides livres e vitamina A) e os não carotenóides (fosfo-lipídios). A superfície da mucosa olfatória oscila em torno de três centímetros quadrados e está separada do restante da cavidade nasal pelo corneto médio, Wageman20. Sendo o corneto médio revestido por uma mucosa extremamente vascularizada e erétil, só permite a passagem de uma pequena quantidade de ar para a região olfatória. O espaço útil por onde o ar inspirado atinge a zona olfatória mede um milímetro de largura por vinte milímetros de comprimento. As reações vasomotoras bruscas ocasionam o fechamento dessa fenda, isolando a mucosa olfatória de um estímulo mais forte ou nocivo. Mc Leod14 fala mesmo de uma verdadeira "pupila olfativa".
CÉLULA OLFATIVA - O epitélio olfatório é formado de células sensoriais engastadas entre as de sustentação. A célula olfatória ou célula de Schulrze é um neurônio bipolar, fusiforme, cuja situação periférica é exclusividade dessa parte do organismo Esta disposição sem similar do ponto de vista anatômico, notável por sua simplicidade, mostra a antiguidade da função olfatória, Gutierrez11. Nos outros órgãos sensoriais a célula sensorial é distinta do neurônio sensitivo. Essas células olfativas apresentam dois prolongamentos: o distal e o proximal. O prolongamento distal, na espessura do epitélio, aflora à superfície entre as células de sustentação e termina por um alargamento hemisférico chamado vesícula olfatória. Essa vesícula é coberta de cílios rígidos, curtos, e envolvidos pelo muco olfatório que banha em grande quantidade este epitélio. No microscópio eletrônico, uma vesícula olfatória se apresenta como um tufo de cílios que mede quinhentos angstroms de diâmetro e 0,5 micra de comprimento. O número de cílios varia em cada espécie animal, Graziadei10 e De Lorenzo5. No homem, são de aproximadamente, segundo Bloom e Engstron, 1 000 por célula e cada um mede cerca de 0,1 micra de espessura e 100 a 200 mitras de comprimento. Ao contrário dos cílios da mucosa nasal respiratória, os cílios da mucosa olfatória são imóveis. Pela existência desses cílios, a área do receptor olfatório aumenta consideravelmente, e segundo Mc Leod14 é multiplicada por 100, ocupando portanto 300 centímetros quadrados. O prolongamento proximal da célula inicia um cilindro-eixo amielínico muito fino de 0,1 a 0,2 mitras de diâmetro que depois de atravessar a lâmina crivada do etmoide, termina numa arborização sináptica dentro de um glomerulo do bulbo olfatório, onde ele se relaciona com os dendritos do segundo neurônio. No homem há mais ou menos 200 milhões de receptores olfatórios e outros tantos de fibras olfatórias.
CÉLULAS DE SUSTENTAÇÃO - Ocupam toda a altura do epitélio, são esquematicamente cilíndricas e envolvem a base das células sensoriais havendo interrelação entre elas; Jahn12, Logan Turnerla. A sua função é ainda desconhecida, sendo-lhe atribuída a função secretora, a de limpeza dos cílios e a ação fagocitária.
CÉLULAS BASAIS - Formam na face profunda do epitélio uma camada única logo acima do corion. Sua função seria a de substituir as células de sustentação.
CORION OU TÚNICA PRÓPRIA - Separada do epitélio por uma membrana basilar homogênea encontramos a túnica própria, que é composta de um arcabouço conjuntivo feito de fibras colágenas e reticulócitos, repousando sobre o esqueleto ósseo nasal. O elemento primordial do corion é a glândula túbulo-acinosa de Bowman que sintetiza uma secreção mista sero-mucosa a qual é eliminada na superfície do epitélio por um canal excretor sinuoso. A composição química dessa secreção varia de acordo com o estímulo olfatório. Para Fortunato e Nicolini8 a secreção glandular além da função de limpeza da mucosa, influenciaria também no processo olfatório. O fluido que cobre esta região não é somente oriundo das glândulas de Bowman, parecendo para Doueke que o lençol de muco, originado da mucosa respiratória do nariz e dos seios paranasais, está também presente. Para a maioria dos autores, as células olfatórias não têm capacidade de regeneração. Van Scerpenberg18 põe em dúvida essa asserção, concebendo a possibilidade de regeneração do epitélio olfatório. Ele considera que as células das glândulas de Bowman podem transformar-se em células de sustentação c. em verdadeiras células olfatórias.
NERVO OLFATÓRIO - Está formado pela reunião dos prolongamentos proximais das células de Schultze. No ser humano existem, como já dissemos, 200 milhões de células olfatórias e portanto 200 milhões de fibras nervosas amielínicas de 0,1 a 0,2 mitras de diâmetro que, atravessando a lâmina crivada do etmoide, terminam no bulbo olfatório ao nível do glomérulo, fazendo sinápse com os dendritos do segundo neurônio da via olfatória. Não existe, portanto, nenhum plexo de neurônios ao nível da mucosa olfatória, uma vez que as fibras caminham completamente separadas. Adrian2, baseado na relação de um para um entre as células receptoras e os axônios, demonstrou que no bulbo olfatório do coelho existe uma projeção regional da mucosa olfatória. Assim, a região superior da mucosa olfatória se projeta na parte superior, a região inferior corresponde às zonas inferiores do bulbo. O mesmo ocorre com as áreas anteriores e posteriores. Le Gros Clark e Warwick demonstraram que a ablação do bulbo olfatório causa o degeneração retrograda das células receptoras.
RINENCÉFALO - Denomina-se rinencéfalo as porções de cada hemisfério cerebral relacionadas com a recepção, condução e integração das sensações olfatórias, Erhart7. O rinencéfalo não é uma estrutura anatômica delimitada, é um conjunto de formações responsáveis pela discriminação e percepção consciente dos odores. Segundo Gray, o rinencéfalo é constituído pelo bulbo olfatório tracto olfatório, pirâmide olfatória, tubérculo olfatório pré-piriforme, substância olfatória Perfurada anterior, área piriforme olfatória, formação hipocampal olfatória, giro para-terminal olfatório e núcleo habenular. Estas formações são relativamente atrofiadas no homem. Nos animais inferiores, as estruturas rinencefálicas são a parte fundamental do cérebro. Este cérebro inferior rege o comportamento geral, tem memória, conexões com o sistema somativo e vegetativo, visceral, gustativo, visual, com a região diencéfalo - hipotalâmica e desempenha um papel importante na atividade sexual, de agressividade e de defesa. No dizer de Chouard4 "o rinencéfalo, notável por suas vias aferentes e eferentes, pode ser considerado como um verdadeiro cérebro do comportamento." No homem, este papel é desempenhado pelo neopallium (córtex cerebral).
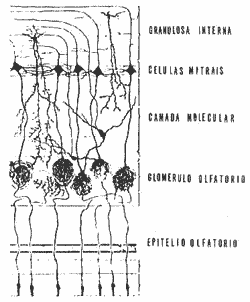
BULBO OLFATÓRIO - É um dos componentes do rinencéfalo, tem a forma ovóide e repousa sobre a lâmina crivada do etmóide sob o lobo frontal. Ele é formado de acordo com o esquema 1, de 6 camadas assim dispostas: fileira olfatória, glomérulos olfatórios, granulosa externa, molecular, células mitrais e granulosa interna. Na superfície do bulbo existem de 20 a 25 mil glomérulos que, como vimos, traduzem a sinápse entre o prolongamento proximal das células de Schultze e o prolongamento protoplasmático dos primeiros neurônios centrais. Esses neurônios compreendem as células mitrais, mais numerosas e importantes, as células em penacho e as granulares, muito abundantes na camada molecular e na granulosa interna, onde constituem curtos neurônios de associação. Ao nível do bulbo olfatório existe uma extraordinária concentração do estímulo nervoso originado da célula olfatória. Segundo Mc Leodla esse estímulo que parte de um dos 2000 milhões de receptores, faz "relais" com um dos 40 mil glomérulos para ser finalmente recolhido por uma das 4 a 5 mil células mitrais. Assim cada célula mitral se relaciona com 4 a 5 glomérulos e cada glomérulo se relaciona com 18 mil células receptoras.
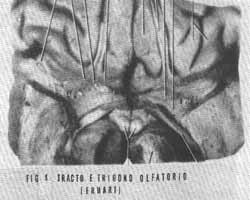
TRACTO OLFATÓRIO - O tracto olfatório, cinta ou estria, figura 2, percorre o sulco olfatório localizado no lobo frontal. É formado pelos axônios centrípetos mielínicos das células mitrais, e das células em penacho e também de fibras provenientes do bulbo olfatório contralateral que chegam a ele cruzando a comissura anterior. Após um trajeto aproximado de 2 centímetros, junto à substância perfurada anterior, este tracto se bifurca nas estrias olfatórias lateral e medial, demarcando o trígono olfatório, figura 1.
A estria olfatória lateral vai em direção ao suco lateral, alcança o córtex da insula na área piriforme, nele penetrando, emitindo colaterais, e termina no corpo do núcleo amigdalóide, Erhart7. A estria olfatória medial dirige-se, apoiada no giro olfatório medial, à área subcalosa e ao pendúculo do corpo caloso. A pequena eminência localizada na substância perfurada anterior, atrás do trígono olfatório, chama-se tubérculo olfatório, sendo considerada como formação cortical rudimentar. Dessas estruturas enumeradas saem outras fibras nervosas cujo trajeto ainda não está perfeitamente esclarecido, mas que parece relacionado com os vários reflexos olfatórios, nada tendo a ver com a percepção dos odores. Zilrtorff21 afirma que ao estudarmos o sentido do olfato não podemos esquecer que vastas porções do rinencéfalo não têm relação com o mesmo, e que os centros primários olfatórios compreendem somente o uncus e a parte anterior do giro do hipocampo. A córtex orbitária, na vizinhança do uncus do hipocampo, e também a área de projeção das sensações gustativas, ZjIStorff22. Allison apud Douek, destruindo a área pré-piriforme de coelhos, demonstrou que o sentido do olfato, tal qual os outros sentidos, é finalmente representado na iso-córtex. Pode-se também afirmar que o sentido do gosto tem uma localização iso-cortical na região sensacão somática da líneua. As áreas gustatórias e olfatórias estão muito próximas entre si e da região onde, por estímulos elétricos, se induz a salivação e a mastigação.
Desta forma procura-se materializar as relações existentes entre gustação e olfato. Subjetivamente, as sensações olfatórias e gustatórias estão completamente integradas. Esse fato é facilmente constatado nos resfriados fortes, durante os quais o indivíduo tem um hiposmia temporária e os alimentos perdem o sabor característico. O paladar é responsável pelos 4 sabores essenciais (amargo, azedo, doce e salgado), as outras variações do gosto são essencialmente olfatórias. Numa comparação, o gosto traduziria as 7 notas fundamentais do piano e o olfato representaria o mundo infinito dos harmônicos. Podemos afirmar que a córtex olfatória, localizada no lobo frontal e temporal, está unida indiretamente às múltiplas zonas corticais, subcorticais e ao rinencéfalo, Adeg1. Assim os impulsos olfatórios oriundos da área piriforme atingem o hipocampo e o giro denteado, de onde fibras eferentes os levam, via alveus, fimbria e fornix, aos núcleos mamilares e à formação reticular do tronco encefálico. Fibras do corpo mamilar se conectam com o núcleo talâmico anterior e com o giro singular. Via "stria terminalis", fibras nervosas chegam ao hipotalamo, núcleos septal e pré ótico vindos do complexo amigdalóide, haveria portanto duas projeções principais de fibras:
1- Terminação cortical principalmente na área pré piriforme, mas também na parte dorsal e externa do núcleo olfatório anterior do tubérculo olfatório e na área córtico medial da amígdala.
2- Terminação sub-cortical na "stria terminalis" e no grupo central amigdalóide. Essa disposição anatômica pode significar que há dois níveis de atividades olfatória:
(a) uma discriminação sofisticada associada em reações de inteligência.
(b) uma discriminação trivial associada com reações instintivas, Douek6.
Todavia, este relacionamento está longe de ser perfeitamente esclarecido.
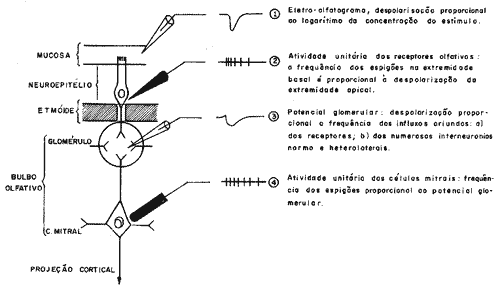
ELETROFISIOLOGIA - As moléculas odoríferas atingindo o receptor olfatório geram potencial elétrico monofásico negativo, único, durante alguns segundos, cuja voltagem aproxima-se de 10 milivolts. Ottoronl16 foi quem obteve pela primeira vez este registro e o denominou de eletro-olfatograma. Quando o microeletródio era colocado numa célula da região respiratória não se registrava variação do potencial elétrico. Ottoson ainda demonstrou que a resposta elétrica é obtida quando o microeletródio (1 micra) é colocado nas células receptoras propriamente ditas e não nas fibras nervosas. Isto, porque a resposta permanece invariável depois do bloqueio anestésico com cocaína, a qual, como sabemos, não afeta as células sensitivas e somente as fibras nervosas. O comportamento da resposta elétrica não é constante, variando pouco com a natureza da substância odorífera estimulante. .Ela é mais durável quando a substância é mais lipotrópica e mais breve quando a substância odorífera é mais hidrosolúvel, Mc Leod14. Do estudo comparativo de vários olfatógramas Mc Leod concluiu que se pode decompô-los em 3 partes: uma deflexão eletronegativa breve, mas extensa, uma deflexão eletropositiva média e uma deflexão eletronegativa longa, mas pouco intensa. De acordo com o esquema 2 relacionando estes diferentes achados, podemos esboçar a atividade elétrica do órgão olfativo.
Esquema 2 - Esquema dos diferentes pontos onde se pode medir a variação de potencial (segundo Mac Leod).
O primeiro fato é a atividade elétrica do receptor em resposta ao sinal químico. Após a chegada de uma mistura de ar carregada de um vapor odorífero, e de uma latência de 0,1 a 0,2 segundos, aparece na superfície do neuroepitélio uma despolarização de amplitude de alguns milivolts que atinge rapidamente seu valor máximo e desaparece em alguns segundos. Em seguida, o estímulo se transmite ao primeiro neurônio em uma freqüência de "espigão" proporcional à despolarização do receptor de onde ele provém, Wrightl9. O sistema de transmissão da informação é, pois, um sistema clássico por modulação de freqüência. O estímulo químico engendra uma despolarização que, segundo Ottoronl5, é proporcional ao logarítimo da intensidade do estímulo o qual se transmite sob a forma de um influxo aferente. Um sistema análogo se encontra na junção do segundo neurônio. Um influxo aferente libera ao nível da sinápse glomerular, um intermediário químico que determina, à altura dos dendritos das células mitrais, uma despolarização que se transmite por modulação de freqüência. Este estímulo é registrado sob a forma de "espigão" nas células mitrais. Adrian2 registrando os potenciais de atividades do bulbo olfativo pôde constatar dois fatos: 1 - quando o eletródio é implantado na zona das células mitrais, excitando o órgão receptor olfatório, se obtém uma mistura de potenciais em "espigão' que corresponde ao potencial de ação de uma célula mitral; 2 - quando o eletródio é colocado na superfície do bulbo olfatório, com a finalidade de registrar sua atividade global, obtém-se "ondas induzidas", oscilações quase sinusóides de 30 a 60 ciclos por segundo. Gesteland9, trabalhando com microeletródios, registrou a atividade de um só receptor olfatório. Demonstrou que as células sensoriais não são sensíveis a todos os odores e que, de uma maneira geral, cada célula de Schultze responde mais ou menos a 60% dos estímulos que lhe são propostos sem evidenciar nenhuma especialização ou sistematização, que segundo a natureza química dos odores, quer pela repartição topográfica dessas células na mucosa olfatória. Parece, então, que esta distribuição aleatória dos receptores olfatórios seja a solução encontrada pela natureza para perceber os odores, mesmo os mais tênues.
A partir do segundo neurônio, de acordo com Chouard4, os registros elétricos tornam-se muito mais fracos e perdem sua especificidade. Pelo jogo dos sistemas inibidores e facilitadores, tudo se passa como se o influxo nervoso se difundisse de segmento em segmento sem que fosse possível segui-lo ou identificá-lo com certeza. Isso explica a dificuldade da interpretação dos achados eletrofisiológicos ao nível do rinencéfalo. Como a experimentação no ser humano não é possível, o mecanismo íntimo da olfação é ainda pouco conhecido, havendo uma distância muito grande nas estruturas anatômicas centrais dos animais em relação ao homem, o mesmo não acontecendo quando essas experiências são realizadas ao nível do epitélio olfativo e do primeiro neurônio, cujas formações histológicas são análogas em ambos. O que conhecemos de concreto deve-se a Penfield e Jasper, Apud Chouard4 ao demonstrarem que só a estimulação elétrica do bulbo olfatório e do uncus do hipocampo é capaz de ocasionar, no homem, a percepção de um odor cuja natureza varia segundo o ponto estimulado em campo limitado (odores forres, desagradáveis), sem que uma topografia sensorial possa ser seguramente estabelecida. Outros autores demonstraram que a estimulação elétrica direta do núcleo amigdalino gera uma crise olfatória acompanhada de uma sensação de medo, despersonalização e delírio alucinatório, colocando em evidência o relacionamento entre a percepção olfatória e os mecanismos do comportamento. Esses fatos foram suscitados por Papez há 20 anos atrás, quando afirmou que o denominado "cérebro-olfatório" estava mais relacionado com o comportamento afetivo do que o olfato. Passouant17, baseado nos trabalhos de Jackson em 1888 e Gowers em 1881, relacionou as implicações existentes entre a epilepsia e o olfato, equacionando o papel da aura olfatória da crise uncinada na facilitação ou inibição de uma crise epiléptica, bem como as modificações do sentido do olfato nos epilépticos.
Concluindo, vemos como são complexas e ainda pouco conhecidas as estruturas e conexões do rinencéfalo. Verificamos que a integração dos impulsos olfatórios e de outras funções viscerais não podem ser dissociadas ou relacionadas somente com o encéfalo. Lembramos, bem a propósito, as palavras de Proetz, "para a exploração do sentido do olfato existem dois obstáculos principais: a simplicidade do órgão terminal e a complexidade do órgão central."
Summary:
The monography, presented in theree main chapters histology of the olfactory mucosa, anatomy of the pathways and cortical centers and eletrophisyology describes the modern concepts on the subject.
Bibliografia
1. Adey, W. R.: Higher olfactory centres. Taste and Smell in Wertebrates: 357-376 J. A. Churchil. London, 1970.
2. Adrian, E. O.: Sensory Messages and sensation. The response of the olfactory organ to different smell. Acta Physiology Scandinavian, 29, 4-14, 1953.
3. Azemar, E.: Parfums et Amours. La France et ses parfums. 8, 137-144 Janvier, 1965.
4. Chouard, C. H.: Aperçus sur la physiologie de Volfaction. Problèmes attuales D'Oto-Rhino-Laryngologie, 125-162 Maloine 1968.
5. De Lorenzo, A. J.: Eletron microscopy of the olfactory and Gustatory path-aways. Ana. Otol. (Sr. Louis) 69: 410-419, 1960.
6. Douek, E.: the sense of smell and its abnormalities. Churchil, Livingstone, 1974.
7. Erhart, E. A.: Neuranatomia. Athenea Editora S. P. 1968.
8. Fortunato, V. Nicolini, P.: L'olfatto. Cognizioni antiche a recenti. Instituto Farmacoterapico Italiano, Roma. 1958.
9. Gesteland, R. C., Lettvin, J. y., Pitts, W. H. and Rojas, A.: Odor Specificitices of the Frogs olfactory receptora. Zotterman edit. Oxford 1, 19-14, 1963.
10. Graziadei, P. P. C.: The mucous membranes of the nose. The annals of Otology Rhinology and Laryngology. 79: 3 433-442, 1970.
11. Gutierrez, C. A.: Frente a un examen de la olfaction. Oto-Laringo-Lógica. IX: 404-407, 1970.
12. Jahn, F. C. Lares, E. C.: Otorrinolaringologia 378-38. Editorial Científico Médica, Barcelona. 1972.
13. L, Turner's: Diseases of the nose, throat and Ear. 26-27 john Wright and sons Ltda., Bristol, 1961.
14. Mac Leod, P.: Physiologie de Volfaction et Discrimination des odeurs. Le concours Médical. 87, 9: 1384-1394, Janvier. 1965.
15. Ottoson, D.: Elettrical signs of olfactory transducer actions. Taste and Smell in Vertebrates: 343-354, J. A. Churchil, 1970.
16. Ottoson, D. and Shepherd, G. M.: Experiment and concepts in olfactory physiology. Progression Brain Research. 83-138, Zoterman Edit. 1967.
17. Passouant P.: Olfaction et épilepsie. La France et ses parfums, 123-136, avril, 1965.
18. Van Scerpenberg, H. J. C.: The possibility of regeneration of the Olfactory epithelium. Excerpta medica. X World Congress of Otorhinolaryngology. 308, 1973.
19. Wright, R. H.: and Burgess, R. E. Specific physicochemical mechanisms of Olfactory Stimulation. Taste and Smell in Vertebrates. 325-335, J. A. Churchil, London. 1970.
20. Wageman, W. K.: Anatomia, Fisiologia y exploration de las fosas nasales y loa senos paranasales, in Tratado de Otorrinolaringologia. Vol. I, 43-52, Berendes-Zollner. Editorial Científico Médico, Barcelona, 1969.
21. Zilstorff, K.: La Otoneurologia Moderna. Oto-Laringo-Lógica. Vol. IX: 1: 6877, 1968.
22. Zilstorff, K.: El examen del sentido del olfato y el tratamiento de las alteraciones del mismo en la practica, Oto-Laringo-Lógica. Vol. IX: 1: 78-86, 1968
Imprimir: ![]()
All rights reserved - 1933 /
2026
© - Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico Facial